

Tubulin glycylases and glutamylases have distinct functions in stabilization and motility of ependymal cilia
- PMID: 23897886
- PMCID: PMC3734080
- DOI: 10.1083/jcb.201305041
Tubulin glycylases and glutamylases have distinct functions in stabilization and motility of ependymal cilia
Abstract
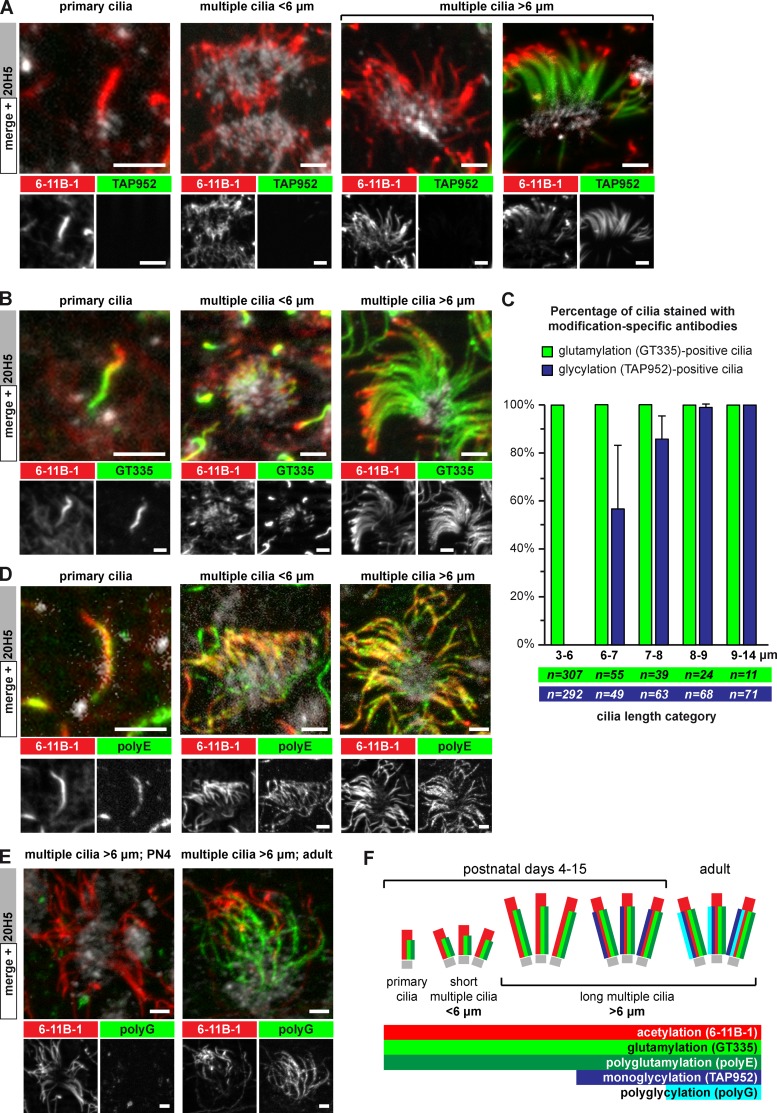
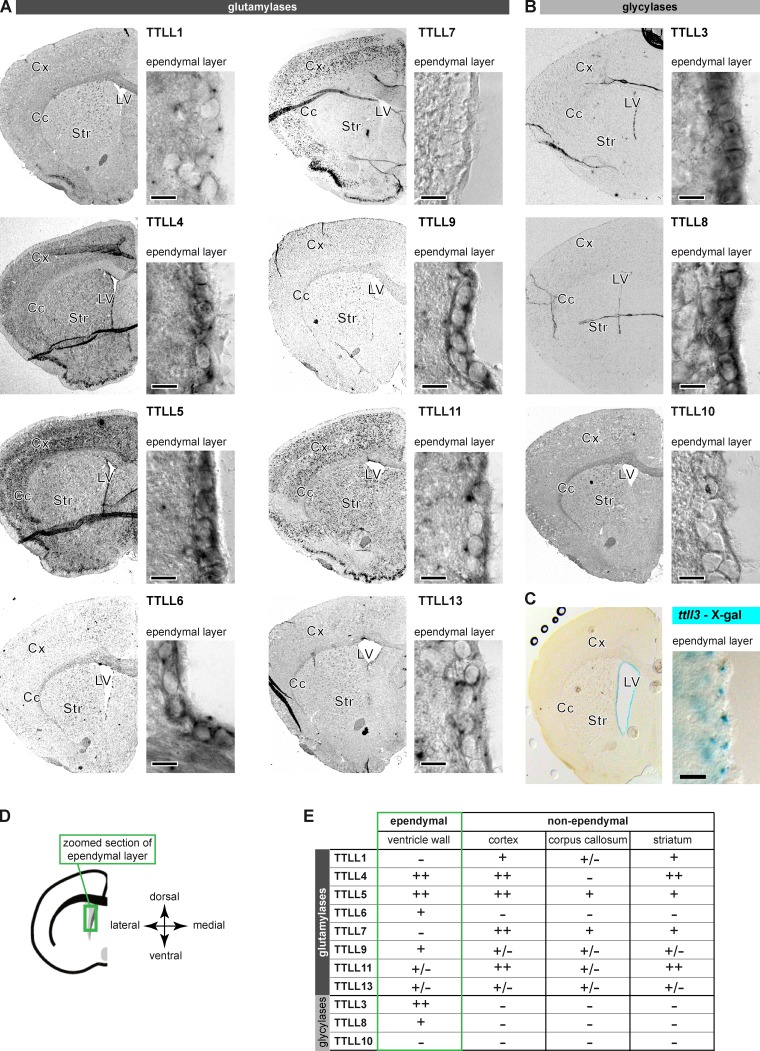
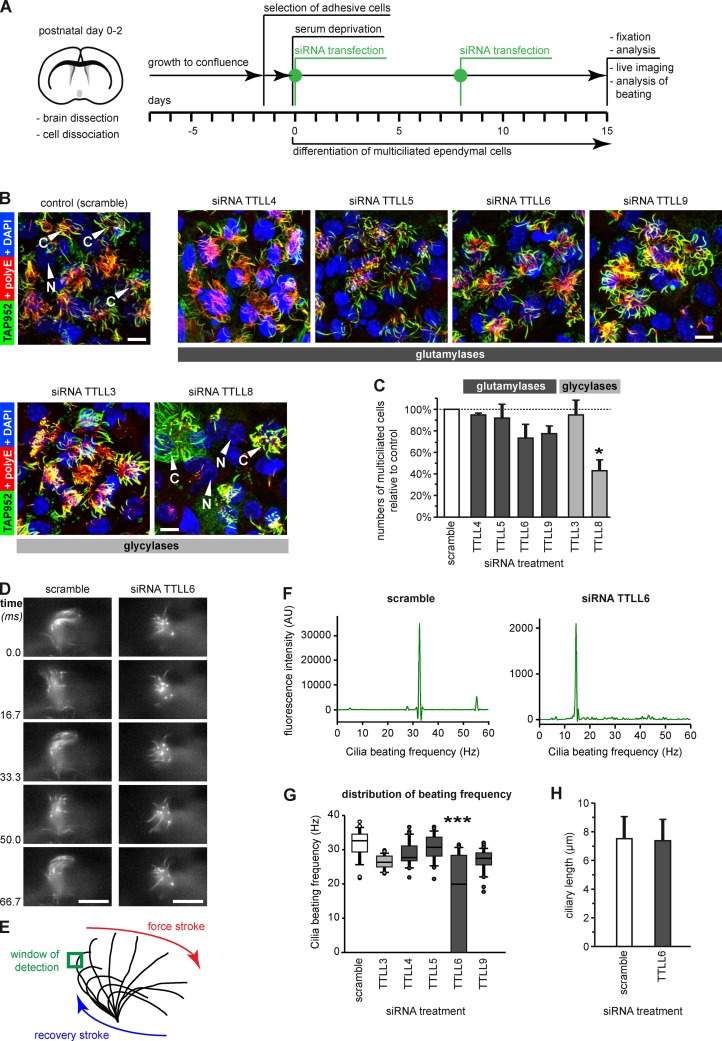
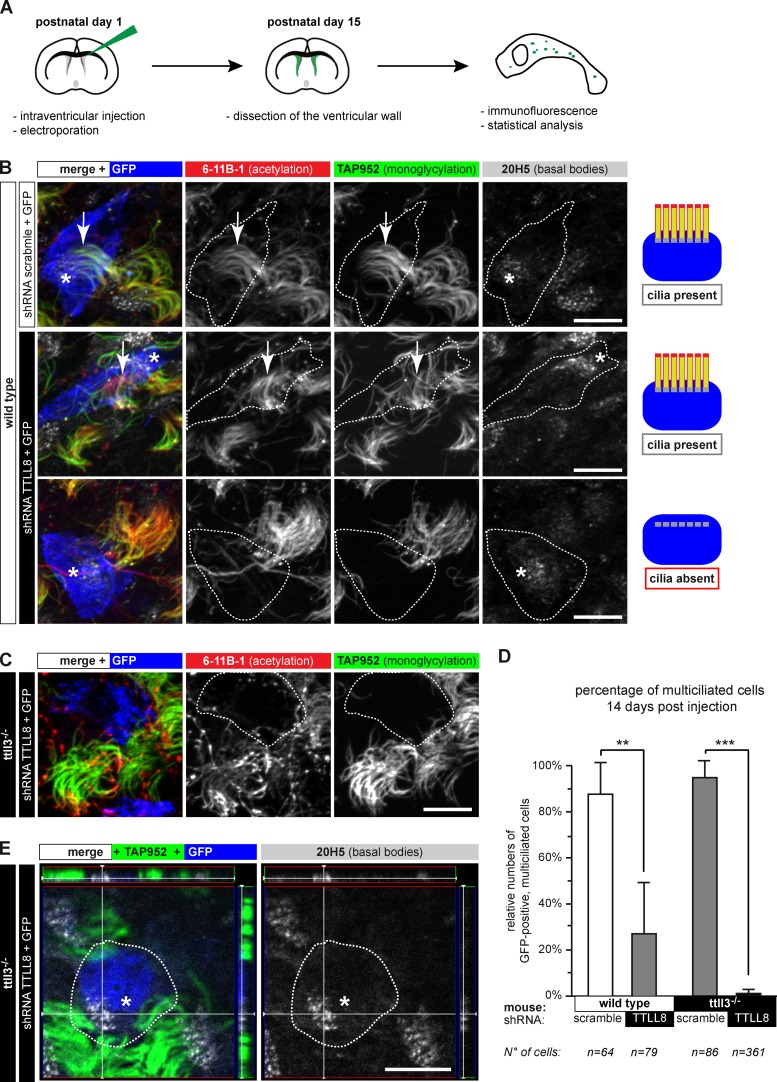
Microtubules are subject to a variety of posttranslational modifications that potentially regulate cytoskeletal functions. Two modifications, glutamylation and glycylation, are highly enriched in the axonemes of most eukaryotes, and might therefore play particularly important roles in cilia and flagella. Here we systematically analyze the dynamics of glutamylation and glycylation in developing mouse ependymal cilia and the expression of the corresponding enzymes in the brain. By systematically screening enzymes of the TTLL family for specific functions in ependymal cilia, we demonstrate that the glycylating enzymes TTLL3 and TTLL8 were required for stability and maintenance of ependymal cilia, whereas the polyglutamylase TTLL6 was necessary for coordinated beating behavior. Our work provides evidence for a functional separation of glutamylating and glycylating enzymes in mammalian ependymal cilia. It further advances the elucidation of the functions of tubulin posttranslational modifications in motile cilia of the mammalian brain and their potential importance in brain development and disease.
Figures

References
-
- Bell P.B., Jr, Safiejko-Mroczka B. 1995. Improved methods for preserving macromolecular structures and visualizing them by fluorescence and scanning electron microscopy. Scanning Microsc. 9:843–857, discussion :858–860 - PubMed
-
- Bré M.H., Redeker V., Quibell M., Darmanaden-Delorme J., Bressac C., Cosson J., Huitorel P., Schmitter J.M., Rossler J., Johnson T., et al. 1996. Axonemal tubulin polyglycylation probed with two monoclonal antibodies: widespread evolutionary distribution, appearance during spermatozoan maturation and possible function in motility. J. Cell Sci. 109:727–738 - PubMed
-
- Carlén M., Meletis K., Göritz C., Darsalia V., Evergren E., Tanigaki K., Amendola M., Barnabé-Heider F., Yeung M.S.Y., Naldini L., et al. 2009. Forebrain ependymal cells are Notch-dependent and generate neuroblasts and astrocytes after stroke. Nat. Neurosci. 12:259–267 10.1038/nn.2268 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
