

JIP1 regulates the directionality of APP axonal transport by coordinating kinesin and dynein motors
- PMID: 23897889
- PMCID: PMC3734084
- DOI: 10.1083/jcb.201302078
JIP1 regulates the directionality of APP axonal transport by coordinating kinesin and dynein motors
Abstract
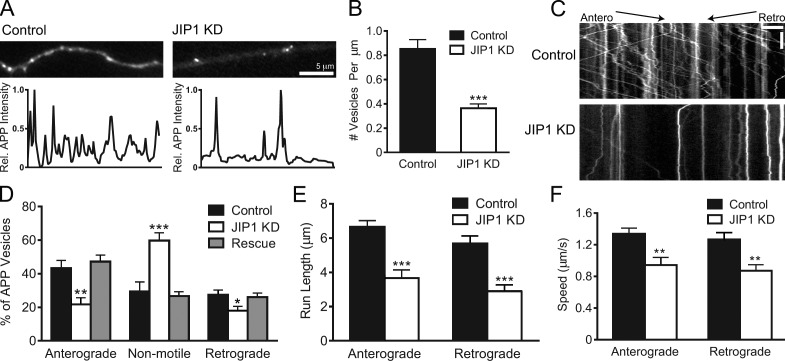
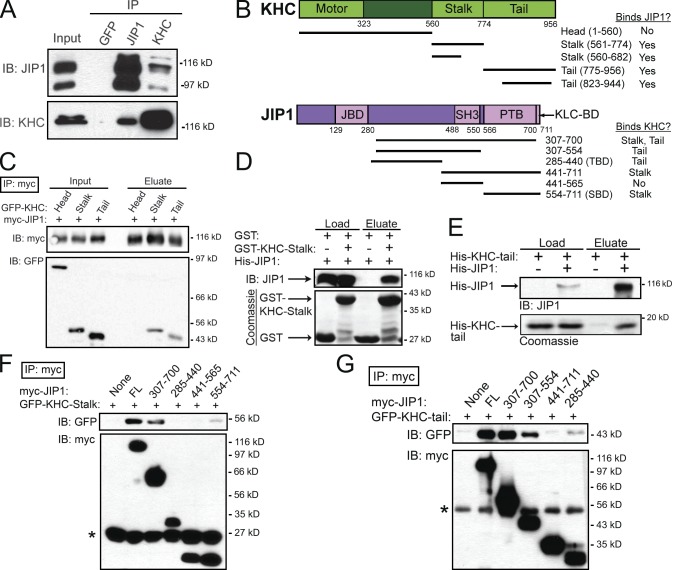
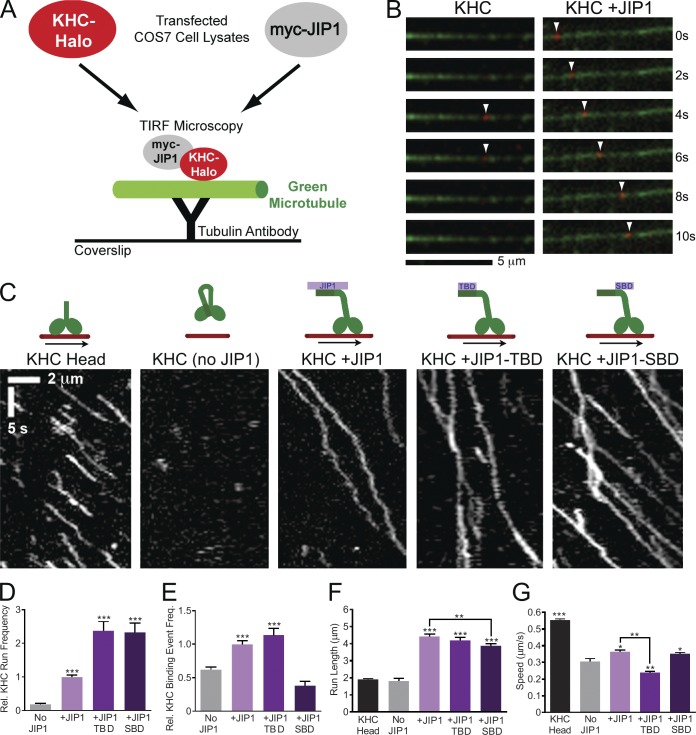
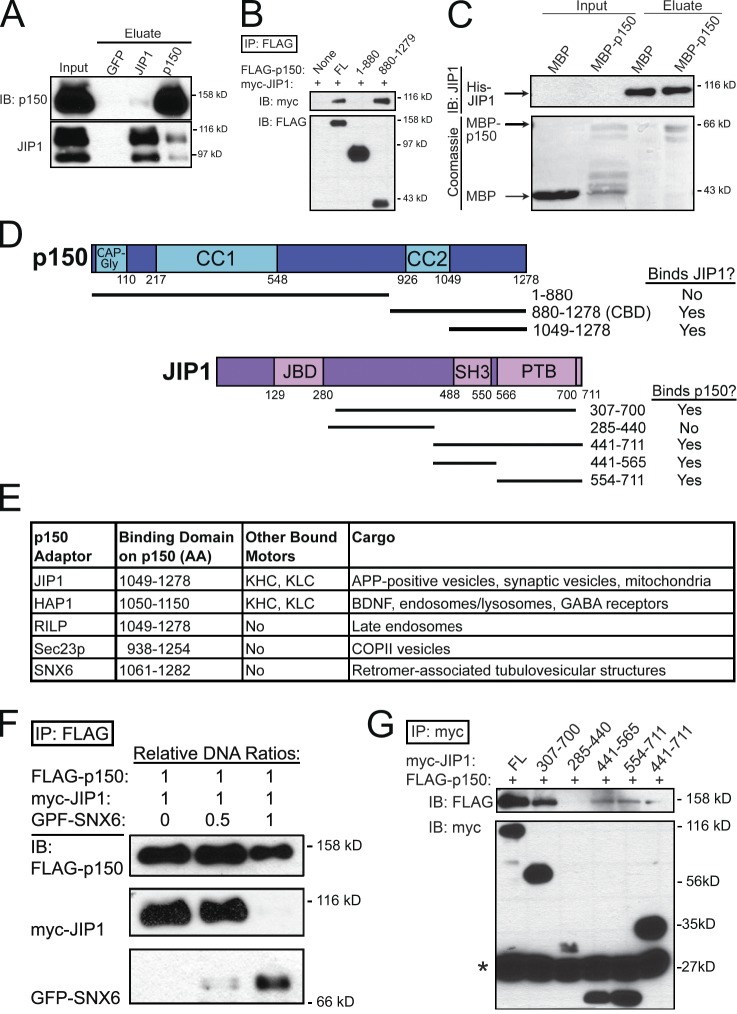
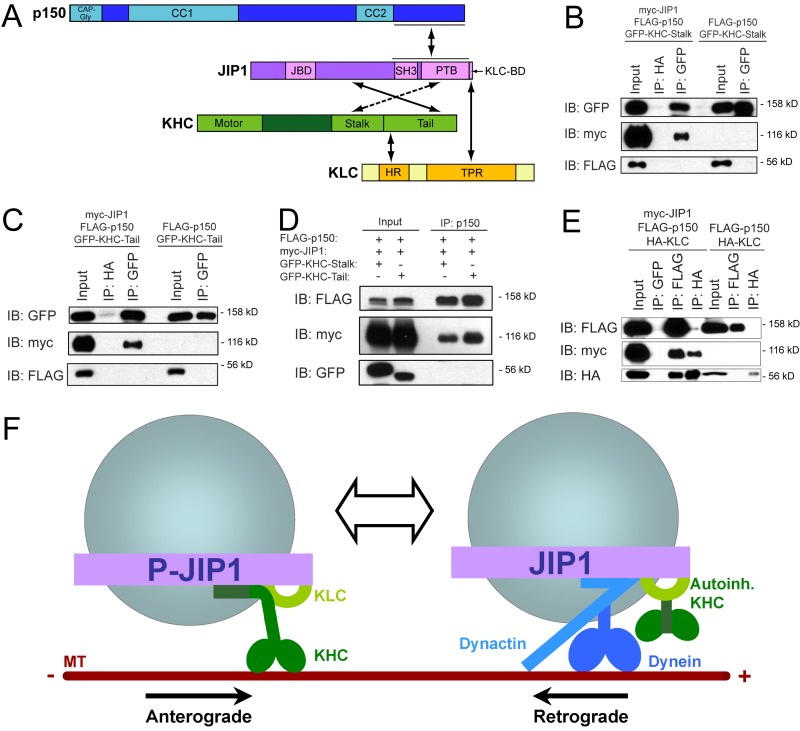
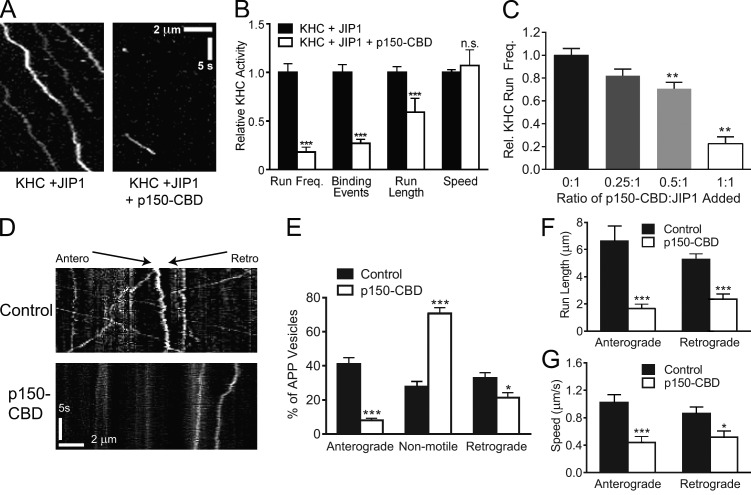
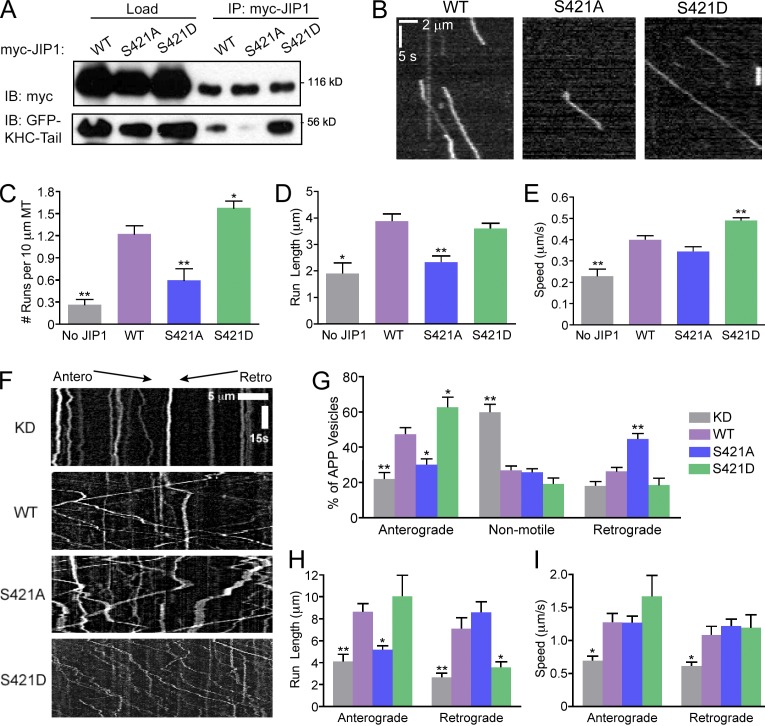
Regulation of the opposing kinesin and dynein motors that drive axonal transport is essential to maintain neuronal homeostasis. Here, we examine coordination of motor activity by the scaffolding protein JNK-interacting protein 1 (JIP1), which we find is required for long-range anterograde and retrograde amyloid precursor protein (APP) motility in axons. We identify novel interactions between JIP1 and kinesin heavy chain (KHC) that relieve KHC autoinhibition, activating motor function in single molecule assays. The direct binding of the dynactin subunit p150(Glued) to JIP1 competitively inhibits KHC activation in vitro and disrupts the transport of APP in neurons. Together, these experiments support a model whereby JIP1 coordinates APP transport by switching between anterograde and retrograde motile complexes. We find that mutations in the JNK-dependent phosphorylation site S421 in JIP1 alter both KHC activation in vitro and the directionality of APP transport in neurons. Thus phosphorylation of S421 of JIP1 serves as a molecular switch to regulate the direction of APP transport in neurons.
Figures
Comment in
-
Slide to the left and slide to the right: motor coordination in neurons.Dev Cell. 2013 Aug 26;26(4):326-8. doi: 10.1016/j.devcel.2013.08.009. Dev Cell. 2013. PMID: 23987508
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
