

Metabolic niche of a prominent sulfate-reducing human gut bacterium
- PMID: 23898195
- PMCID: PMC3746858
- DOI: 10.1073/pnas.1312524110
Metabolic niche of a prominent sulfate-reducing human gut bacterium
Abstract
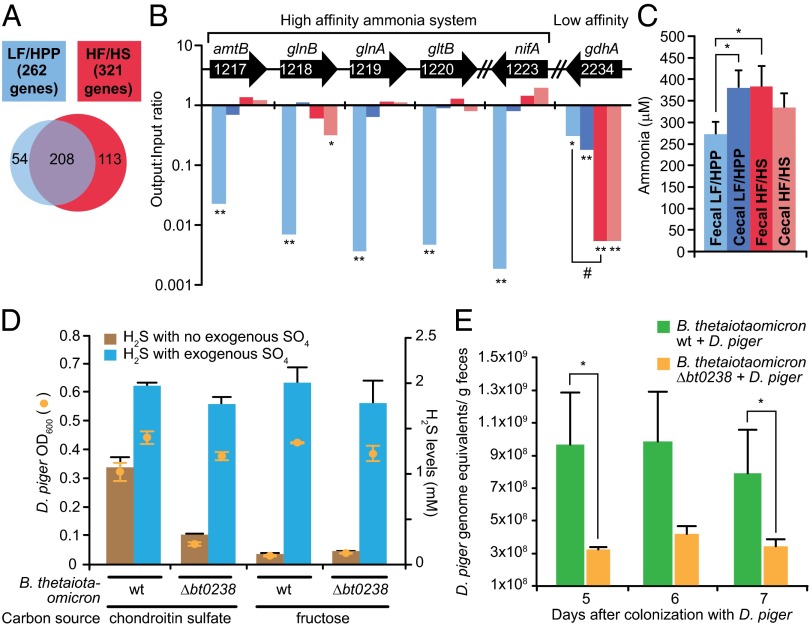
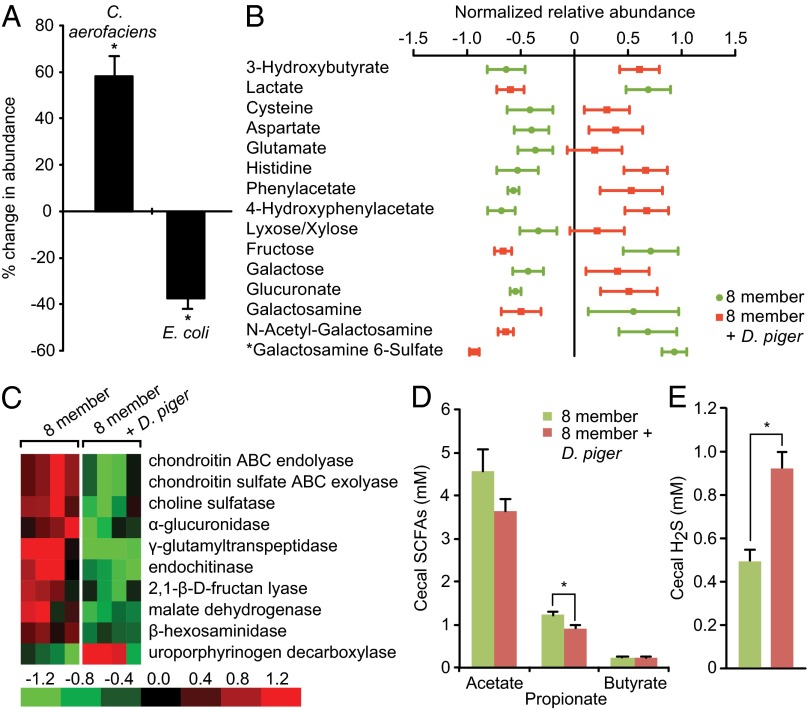
Sulfate-reducing bacteria (SRB) colonize the guts of ∼50% of humans. We used genome-wide transposon mutagenesis and insertion-site sequencing, RNA-Seq, plus mass spectrometry to characterize genetic and environmental factors that impact the niche of Desulfovibrio piger, the most common SRB in a surveyed cohort of healthy US adults. Gnotobiotic mice were colonized with an assemblage of sequenced human gut bacterial species with or without D. piger and fed diets with different levels and types of carbohydrates and sulfur sources. Diet was a major determinant of functions expressed by this artificial nine-member community and of the genes that impact D. piger fitness; the latter includes high- and low-affinity systems for using ammonia, a limiting resource for D. piger in mice consuming a polysaccharide-rich diet. Although genes involved in hydrogen consumption and sulfate reduction are necessary for its colonization, varying dietary-free sulfate levels did not significantly alter levels of D. piger, which can obtain sulfate from the host in part via cross-feeding mediated by Bacteroides-encoded sulfatases. Chondroitin sulfate, a common dietary supplement, increased D. piger and H2S levels without compromising gut barrier integrity. A chondroitin sulfate-supplemented diet together with D. piger impacted the assemblage's substrate utilization preferences, allowing consumption of more reduced carbon sources and increasing the abundance of the H2-producing Actinobacterium, Collinsella aerofaciens. Our findings provide genetic and metabolic details of how this H2-consuming SRB shapes the responses of a microbiota to diet ingredients and a framework for examining how individuals lacking D. piger differ from those who harbor it.
Keywords: artificial human gut microbiota/microbiome; determinants of microbial fitness; hydrogen sulfide; hydrogenotrophs; microbial foodwebs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Wolin MJ, Miller TL. Interactions of microbial populations in cellulose fermentation. Fed Proc. 1983;42(1):109–113. - PubMed
-
- Stams AJ, Plugge CM. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat Rev Microbiol. 2009;7(8):568–577. - PubMed
-
- Cord-Ruwisch R, Seitz HJ, Conrad R. The capacity of hydrogenotrophic anaerobic-bacteria to compete for traces of hydrogen depends on the redox potential of the terminal electron-acceptor. Arch Microbiol. 1988;149(4):350–357.
-
- Scanlan PD, Shanahan F, Marchesi JR. Culture-independent analysis of desulfovibrios in the human distal colon of healthy, colorectal cancer and polypectomized individuals. FEMS Microbiol Ecol. 2009;69(2):213–221. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
