

Longevity effect of IGF-1R(+/-) mutation depends on genetic background-specific receptor activation
- PMID: 23898955
- PMCID: PMC4326867
- DOI: 10.1111/acel.12145
Longevity effect of IGF-1R(+/-) mutation depends on genetic background-specific receptor activation
Abstract
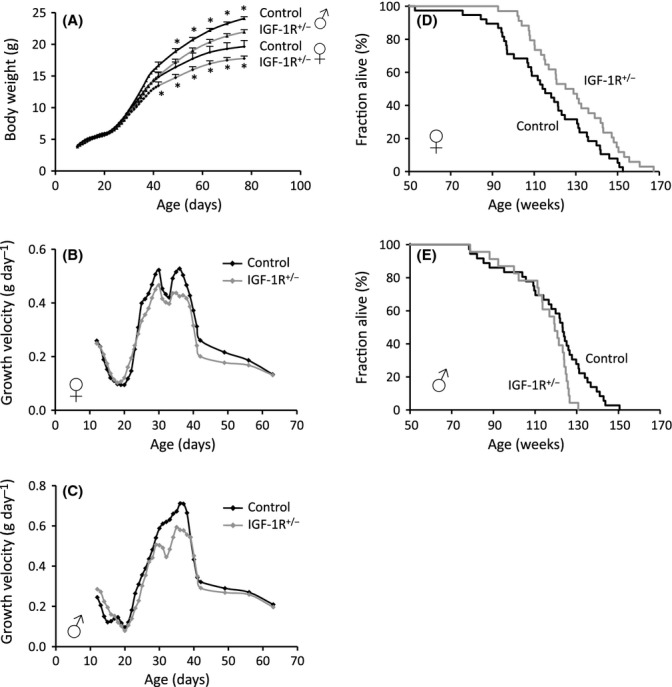
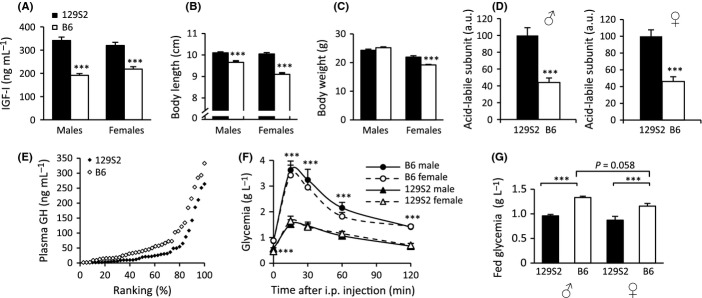
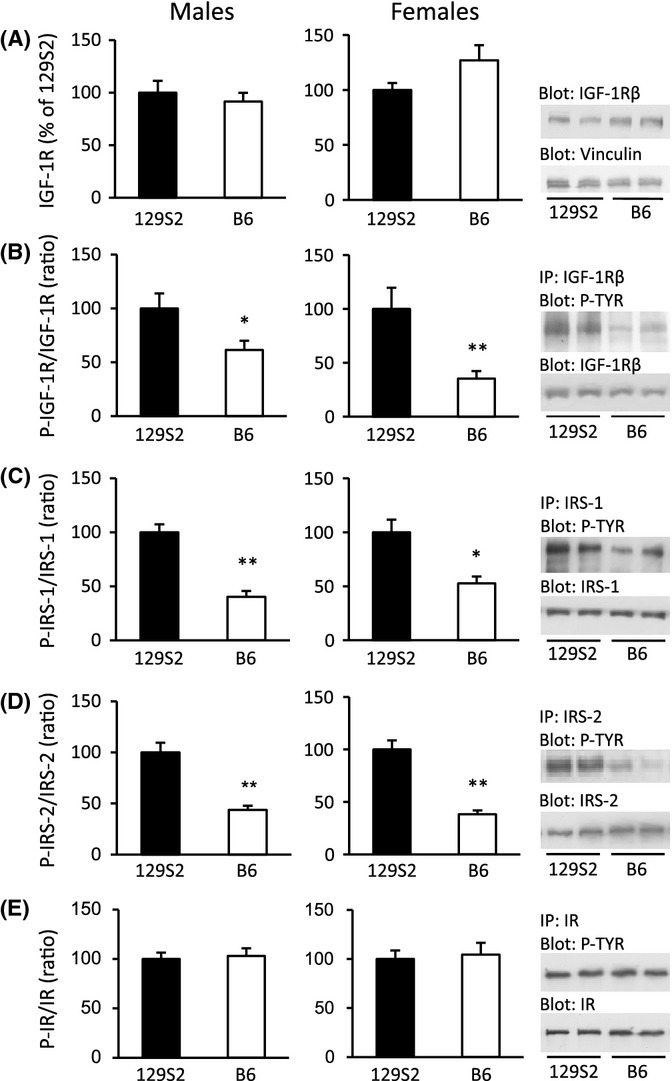
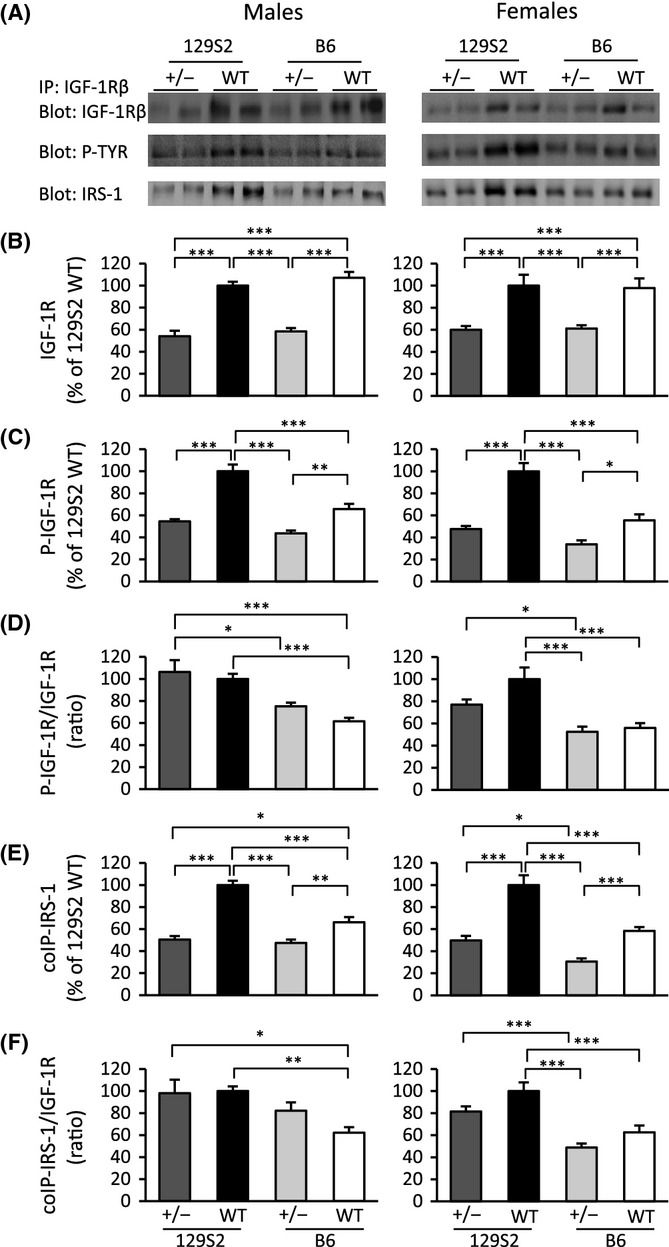
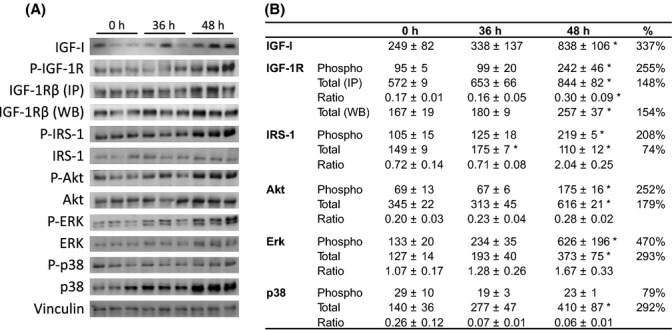
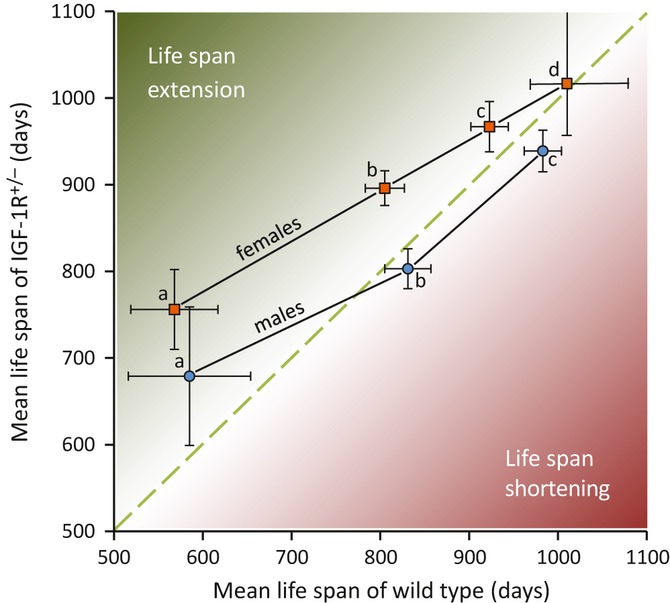
Growth hormone (GH) and insulin-like growth factor (IGF) signaling regulates lifespan in mice. The modulating effects of genetic background gained much attention because it was shown that life-prolonging effects in Snell dwarf and GH receptor knockout vary between mouse strains. We previously reported that heterozygous IGF-1R inactivation (IGF-1R(+/-) ) extends lifespan in female mice on 129/SvPas background, but it remained unclear whether this mutation produces a similar effect in other genetic backgrounds and which molecules possibly modify this effect. Here, we measured the life-prolonging effect of IGF-1R(+/-) mutation in C57BL/6J background and investigated the role of insulin/IGF signaling molecules in strain-dependent differences. We found significant lifespan extension in female IGF-1R(+/-) mutants on C57BL/6J background, but the effect was smaller than in 129/SvPas, suggesting strain-specific penetrance of longevity phenotypes. Comparing GH/IGF pathways between wild-type 129/SvPas and C57BL/6J mice, we found that circulating IGF-I and activation of IGF-1R, IRS-1, and IRS-2 were markedly elevated in 129/SvPas, while activation of IGF pathways was constitutively low in spontaneously long-lived C57BL/6J mice. Importantly, we demonstrated that loss of one IGF-1R allele diminished the level of activated IGF-1R and IRS more profoundly and triggered stronger endocrine feedback in 129/SvPas background than in C57BL/6J. We also revealed that acute oxidative stress entails robust IGF-1R pathway activation, which could account for the fact that IGF-1R(+/-) stress resistance phenotypes are fully penetrant in both backgrounds. Together, these results provide a possible explanation why IGF-1R(+/-) was less efficient in extending lifespan in C57BL/6J compared with 129/SvPas.
Keywords: Genetic background; IGF-I; IRS; gene knockout; lifespan; stress resistance.
© 2013 the Anatomical Society and John Wiley & Sons Ltd.
Figures
References
-
- Ahluwalia A, Clodfelter KH, Waxman DJ. Sexual dimorphism of rat liver gene expression: regulatory role of growth hormone revealed by deoxyribonucleic acid microarray analysis. Mol. Endocrinol. 2004;18:747–760. - PubMed
-
- Blüher M, Kahn B, Kahn CR. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science. 2003;299:572–574. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
