

The circadian clock in cancer development and therapy
- PMID: 23899600
- PMCID: PMC4103166
- DOI: 10.1016/B978-0-12-396971-2.00009-9
The circadian clock in cancer development and therapy
Abstract
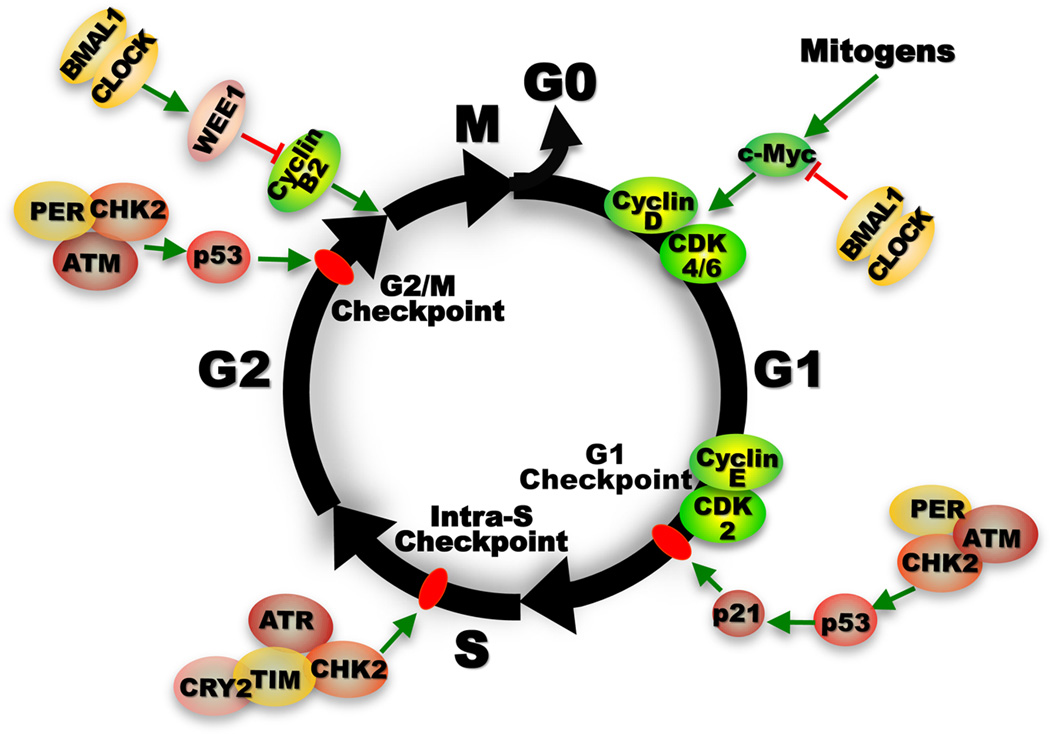
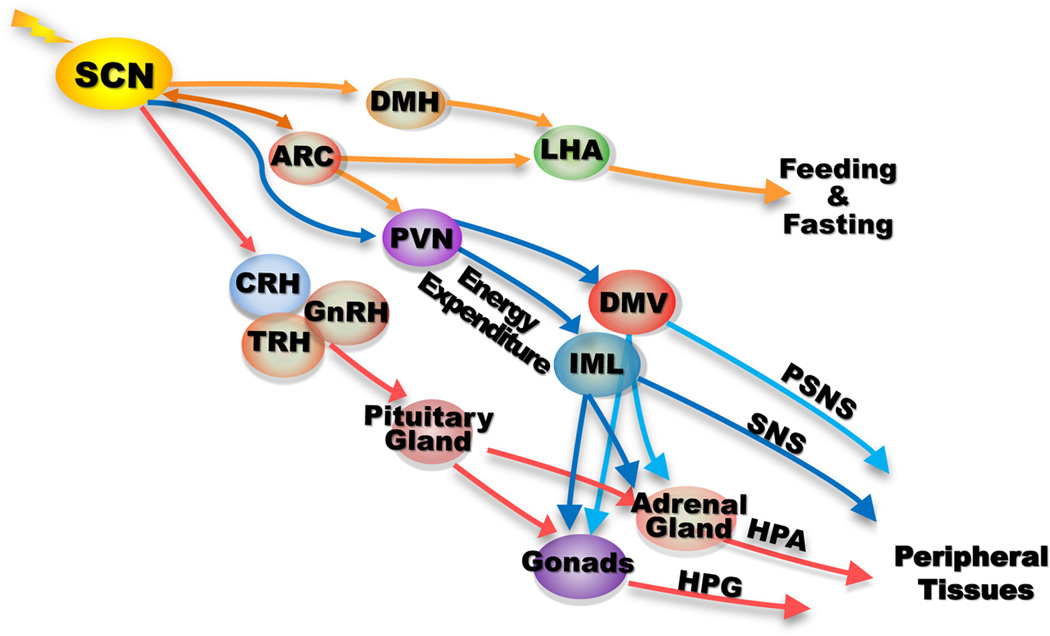
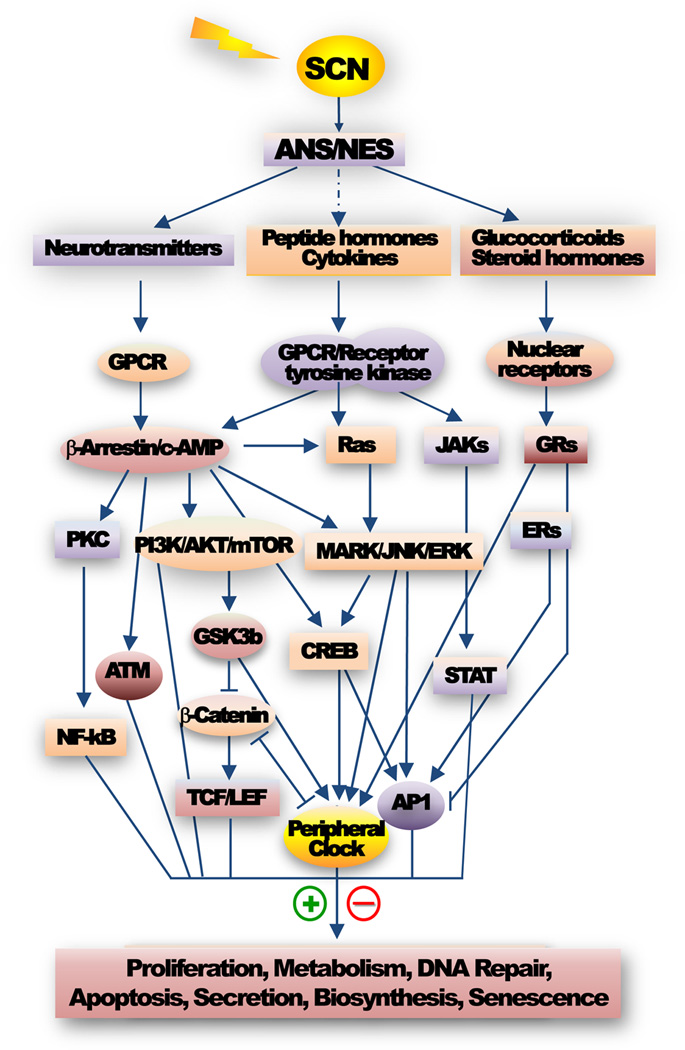
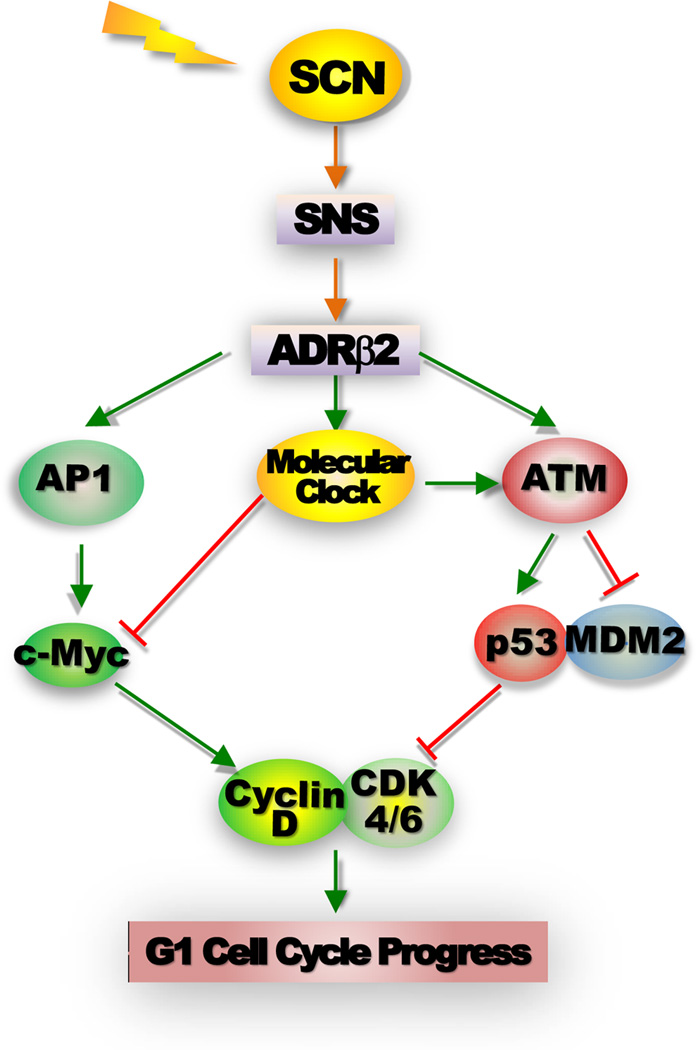
Most aspects of mammalian function display circadian rhythms driven by an endogenous clock. The circadian clock is operated by genes and comprises a central clock in the brain that responds to environmental cues and controls subordinate clocks in peripheral tissues via circadian output pathways. The central and peripheral clocks coordinately generate rhythmic gene expression in a tissue-specific manner in vivo to couple diverse physiological and behavioral processes to periodic changes in the environment. However, with the industrialization of the world, activities that disrupt endogenous homeostasis with external circadian cues have increased. This change in lifestyle has been linked to an increased risk of diseases in all aspects of human health, including cancer. Studies in humans and animal models have revealed that cancer development in vivo is closely associated with the loss of circadian homeostasis in energy balance, immune function, and aging, which are supported by cellular functions important for tumor suppression including cell proliferation, senescence, metabolism, and DNA damage response. The clock controls these cellular functions both locally in cells of peripheral tissues and at the organismal level via extracellular signaling. Thus, the hierarchical mammalian circadian clock provides a unique system to study carcinogenesis as a deregulated physiological process in vivo. The asynchrony between host and malignant tissues in cell proliferation and metabolism also provides new and exciting options for novel anticancer therapies.
Keywords: Aging; Cancer immune surveillance; Cellular senescence; Chronotherapy; Circadian clock; Energy homeostasis; G1 cell cycle progression; Phase shift; Signal transduction; Tumor suppression.
© 2013, Elsevier Inc. All Rights Reserved.
Figures
References
-
- Young MW, Kay SA. Time zones: a comparative genetics of circadian clocks. Nat Rev Genet. 2001 Sep;2(9):702–715. - PubMed
-
- Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002 Aug 29;418(6901):935–941. - PubMed
-
- Buijs RM, Scheer FA, Kreier F, et al. Organization of circadian functions: interaction with the body. Prog Brain Res. 2006;153:341–360. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
