

Two heme-dependent terminal oxidases power Staphylococcus aureus organ-specific colonization of the vertebrate host
- PMID: 23900169
- PMCID: PMC3735196
- DOI: 10.1128/mBio.00241-13
Two heme-dependent terminal oxidases power Staphylococcus aureus organ-specific colonization of the vertebrate host
Abstract
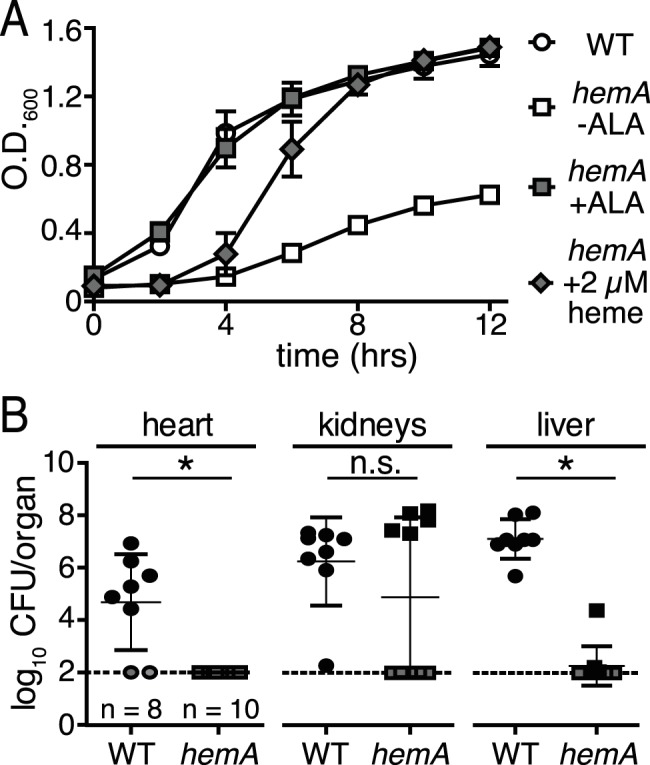
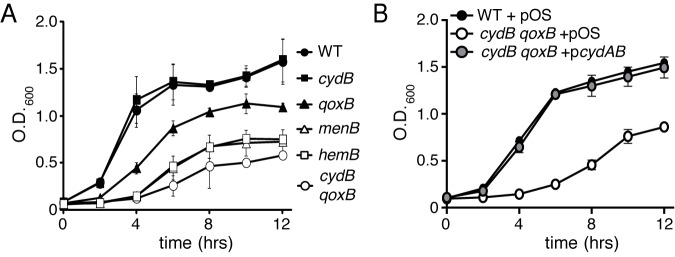
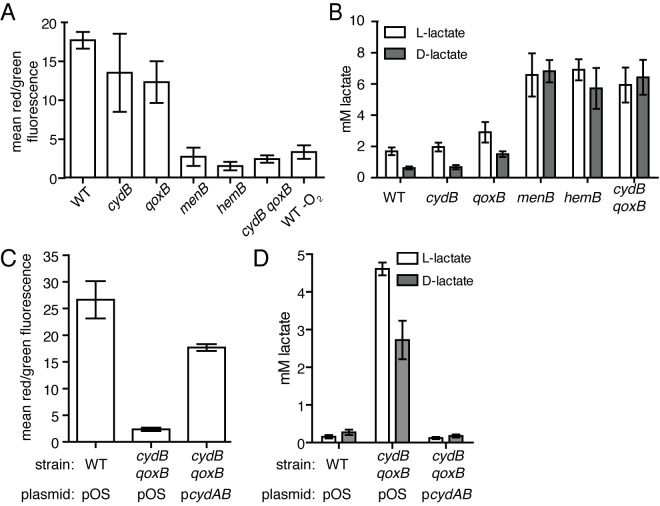
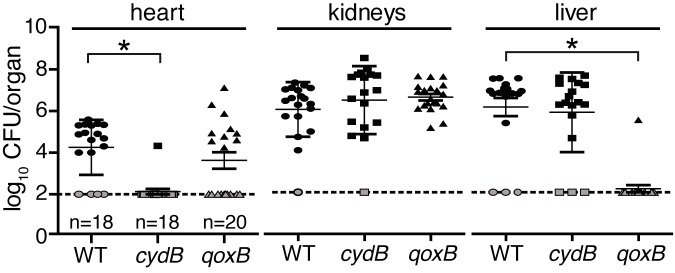
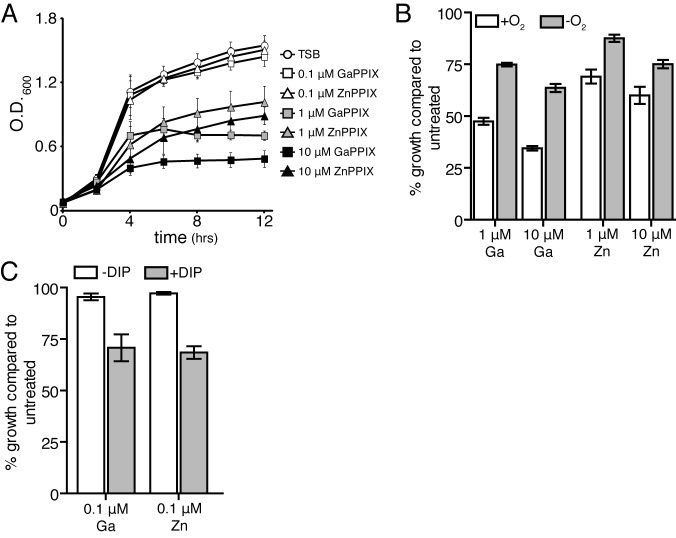
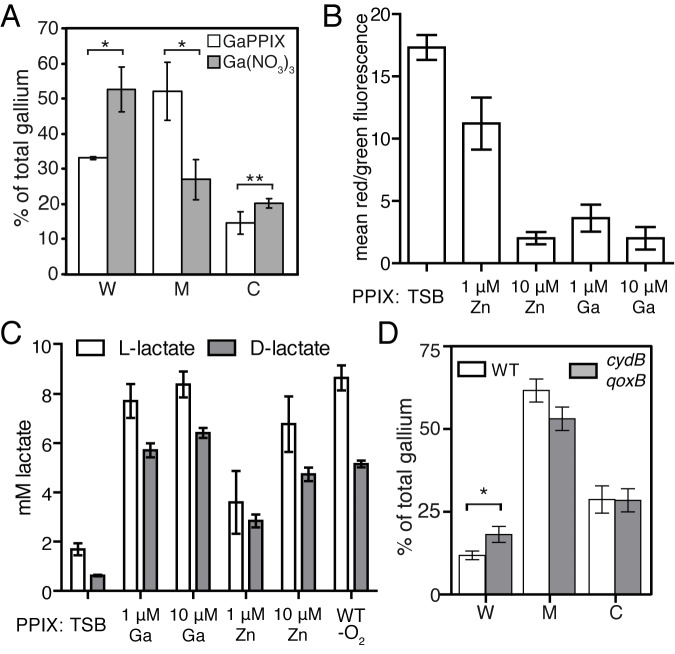
Staphylococcus aureus is a significant cause of infections worldwide and is able to utilize aerobic respiration, anaerobic respiration, or fermentation as the means by which it generates the energy needed for proliferation. Aerobic respiration is supported by heme-dependent terminal oxidases that catalyze the final step of aerobic respiration, the reduction of O2 to H2O. An inability to respire forces bacteria to generate energy via fermentation, resulting in reduced growth. Elucidating the roles of these energy-generating pathways during colonization of the host could uncover attractive therapeutic targets. Consistent with this idea, we report that inhibiting aerobic respiration by inactivating heme biosynthesis significantly impairs the ability of S. aureus to colonize the host. Two heme-dependent terminal oxidases support aerobic respiration of S. aureus, implying that the staphylococcal respiratory chain is branched. Systemic infection with S. aureus mutants limited to a single terminal oxidase results in an organ-specific colonization defect, resulting in reduced bacterial burdens in either the liver or the heart. Finally, inhibition of aerobic respiration can be achieved by exposing S. aureus to noniron heme analogues. These data provide evidence that aerobic respiration plays a major role in S. aureus colonization of the host and that this energy-generating process is a viable therapeutic target.
Importance: Staphylococcus aureus poses a significant threat to public health as antibiotic-resistant isolates of this pathogen continue to emerge. Our understanding of the energy-generating processes that allow S. aureus to proliferate within the host is incomplete. Host-derived heme is the preferred source of nutrient iron during infection; however, S. aureus can synthesize heme de novo and use it to facilitate aerobic respiration. We demonstrate that S. aureus heme biosynthesis powers a branched aerobic respiratory chain composed of two terminal oxidases. The importance of having two terminal oxidases is demonstrated by the finding that each plays an essential role in colonizing distinct organs during systemic infection. Additionally, this process can be targeted by small-molecule heme analogues called noniron protoporphyrins. This study serves to demonstrate that heme biosynthesis supports two terminal oxidases that are required for aerobic respiration and are also essential for S. aureus pathogenesis.
Figures
Comment in
-
Both terminal oxidases contribute to fitness and virulence during organ-specific Staphylococcus aureus colonization.mBio. 2013 Dec 3;4(6):e00976-13. doi: 10.1128/mBio.00976-13. mBio. 2013. PMID: 24302255 Free PMC article.
References
-
- Kuehnert MJ, Kruszon-Moran D, Hill HA, McQuillan G, McAllister SK, Fosheim G, McDougal LK, Chaitram J, Jensen B, Fridkin SK, Killgore G, Tenover FC. 2006. Prevalence of Staphylococcus aureus nasal colonization in the United States, 2001–2002. J. Infect. Dis. 193:172–179 - PubMed
-
- Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, Ray S, Harrison LH, Lynfield R, Dumyati G, Townes JM, Craig AS, Zell ER, Fosheim GE, McDougal LK, Carey RB, Fridkin SK, Active Bacterial Core surveillance (ABCs) MRSA Investigators. 2007. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 298:1763–1771. doi: 10.1001/jama.298.15.1763 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI0169233/AI/NIAID NIH HHS/United States
- R01 AI069233/AI/NIAID NIH HHS/United States
- P30 CA68485/CA/NCI NIH HHS/United States
- T32 HL007751/HL/NHLBI NIH HHS/United States
- AI057157/AI/NIAID NIH HHS/United States
- F32 AI091244/AI/NIAID NIH HHS/United States
- R01 AI073843/AI/NIAID NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- U54 AI057157/AI/NIAID NIH HHS/United States
- F32-AI091244-01/AI/NIAID NIH HHS/United States
- AI073843/AI/NIAID NIH HHS/United States
- L40 AI096516/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
