

The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit
- PMID: 23900286
- PMCID: PMC3771344
- DOI: 10.1038/emboj.2013.157
The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit
Abstract
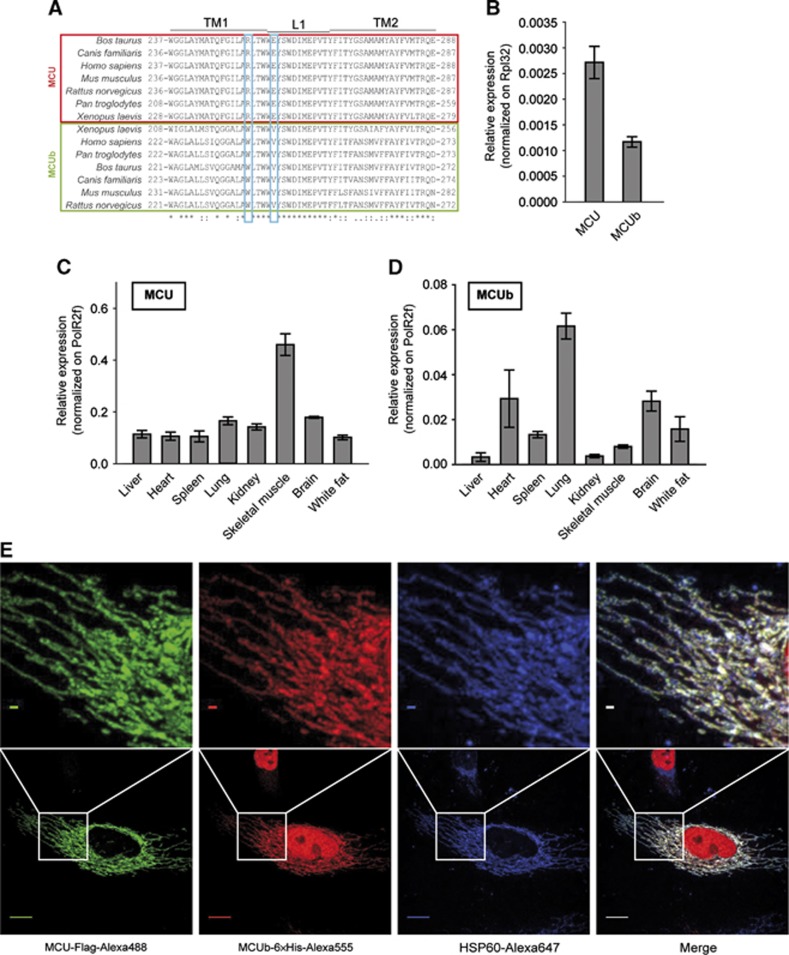
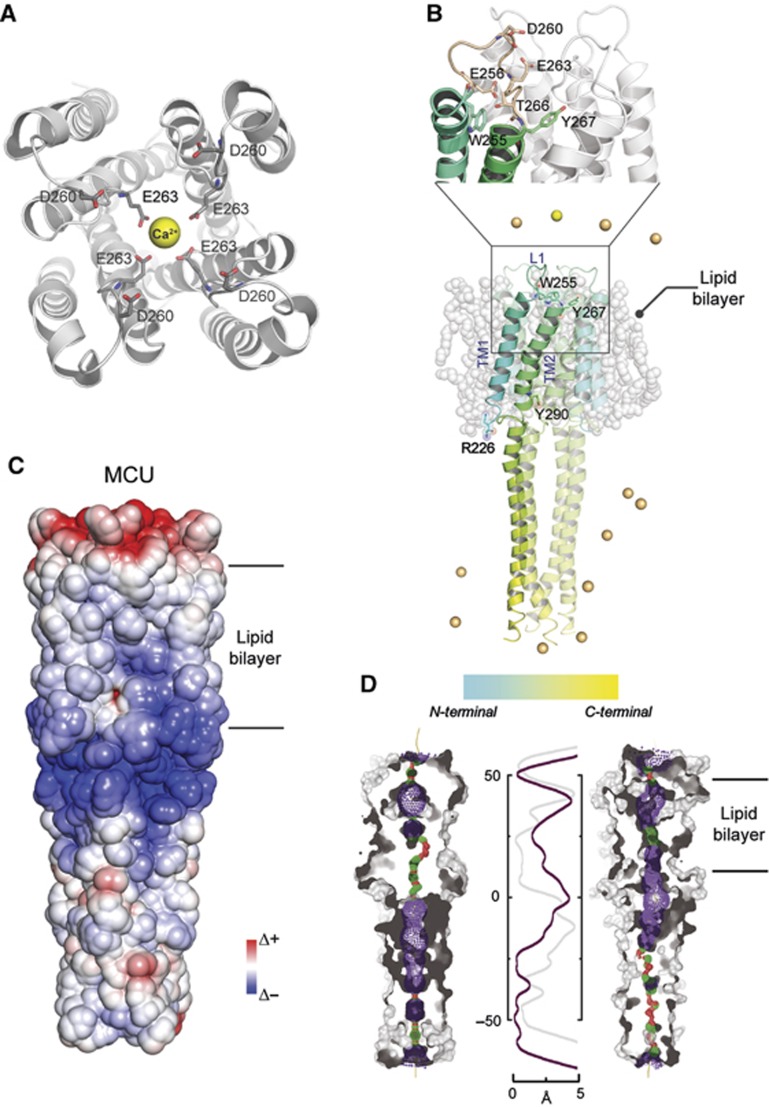
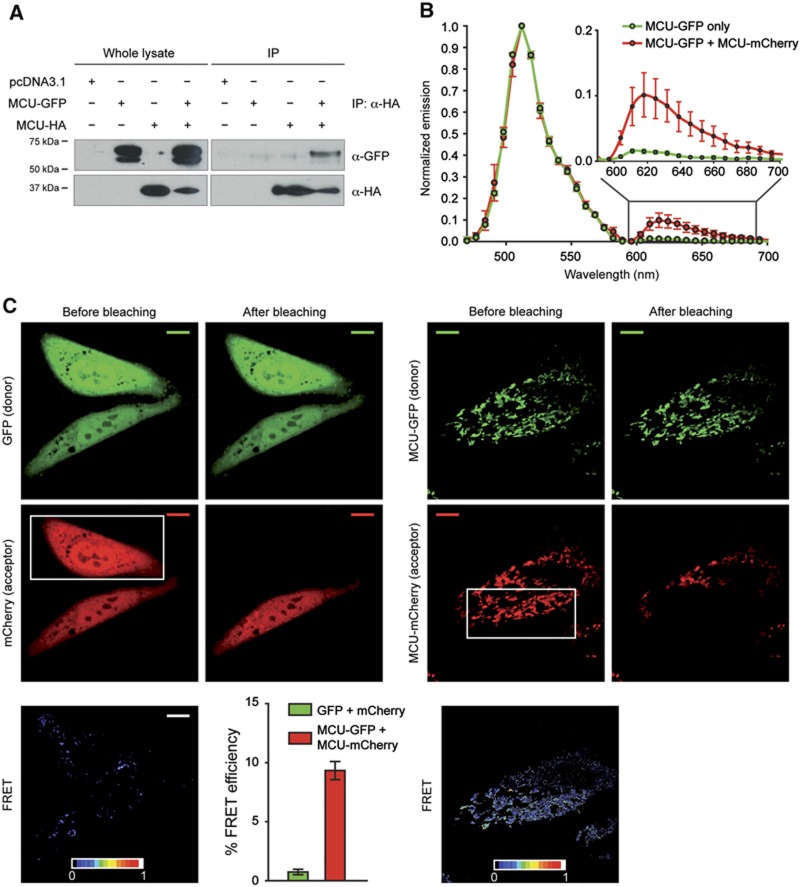
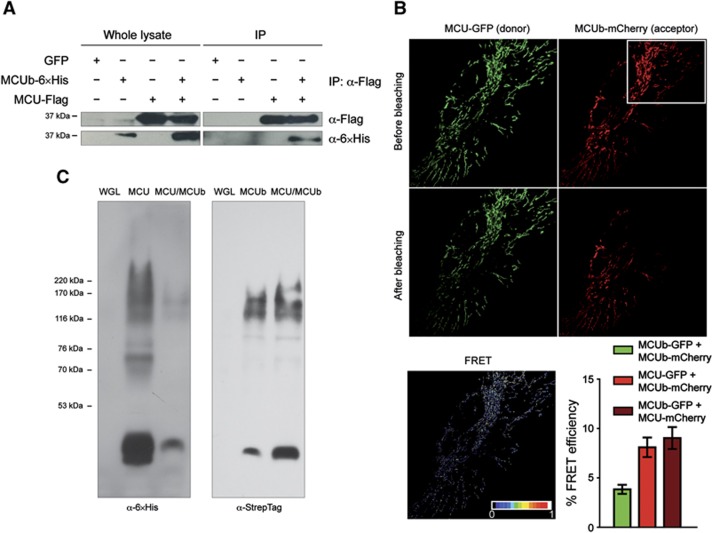
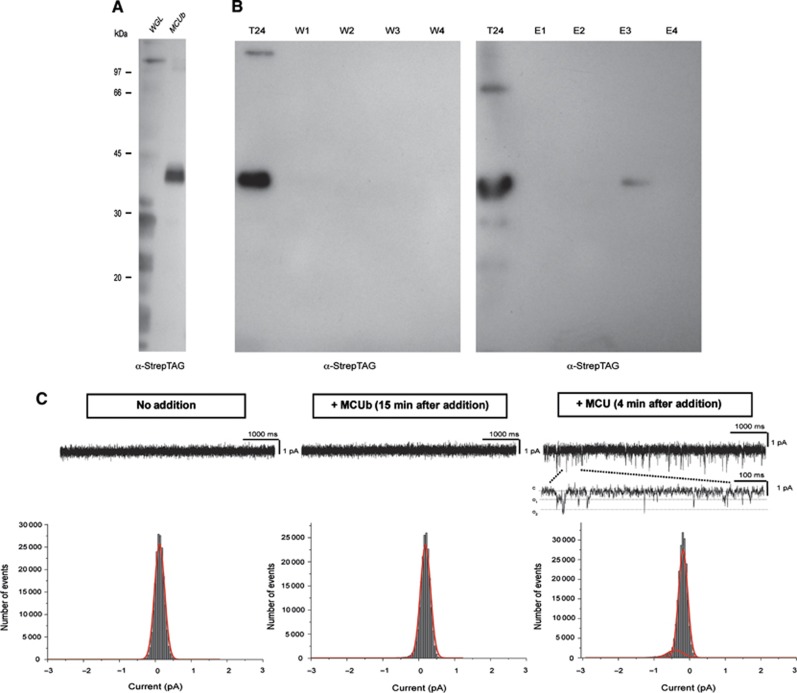
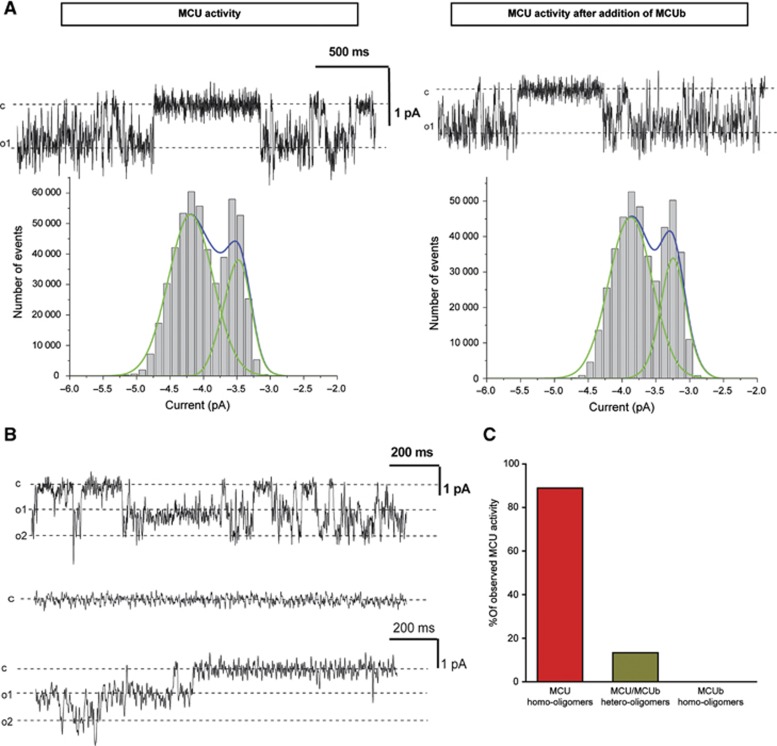
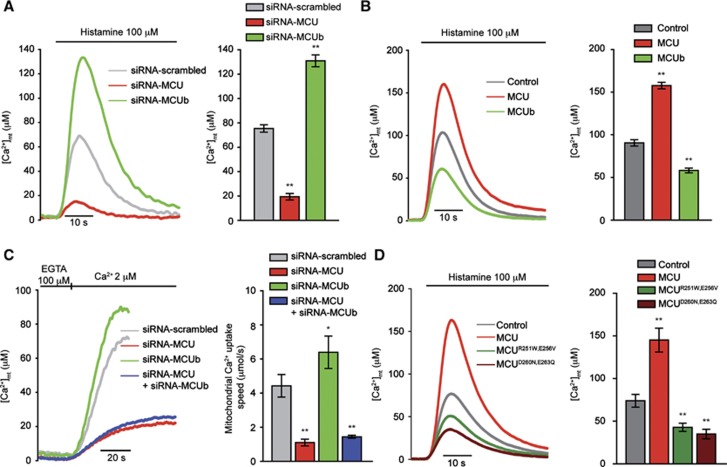
Mitochondrial calcium uniporter (MCU) channel is responsible for Ruthenium Red-sensitive mitochondrial calcium uptake. Here, we demonstrate MCU oligomerization by immunoprecipitation and Förster resonance energy transfer (FRET) and characterize a novel protein (MCUb) with two predicted transmembrane domains, 50% sequence similarity and a different expression profile from MCU. Based on computational modelling, MCUb includes critical amino-acid substitutions in the pore region and indeed MCUb does not form a calcium-permeable channel in planar lipid bilayers. In HeLa cells, MCUb is inserted into the oligomer and exerts a dominant-negative effect, reducing the [Ca(2+)]mt increases evoked by agonist stimulation. Accordingly, in vitro co-expression of MCUb with MCU drastically reduces the probability of observing channel activity in planar lipid bilayer experiments. These data unveil the structural complexity of MCU and demonstrate a novel regulatory mechanism, based on the inclusion of dominant-negative subunits in a multimeric channel, that underlies the fine control of the physiologically and pathologically relevant process of mitochondrial calcium homeostasis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Barel O, Shalev SA, Ofir R, Cohen A, Zlotogora J, Shorer Z, Mazor G, Finer G, Khateeb S, Zilberberg N, Birk OS (2008) Maternally inherited Birk Barel mental retardation dysmorphism syndrome caused by a mutation in the genomically imprinted potassium channel KCNK9. Am J Hum Genet 83: 193–199 - PMC - PubMed
-
- Berridge MJ, Bootman MD, Roderick HL (2003) Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol 4: 517–529 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
