

Polarized light microscopy in reproductive and developmental biology
- PMID: 23901032
- PMCID: PMC4979313
- DOI: 10.1002/mrd.22221
Polarized light microscopy in reproductive and developmental biology
Abstract
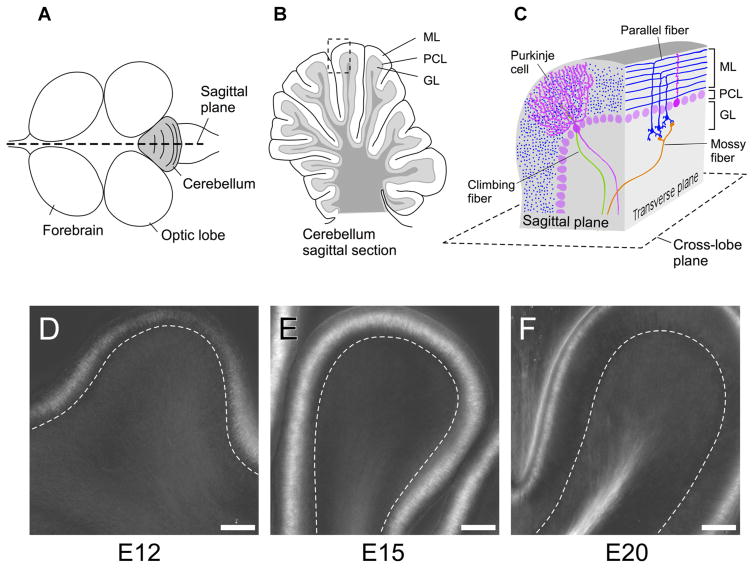
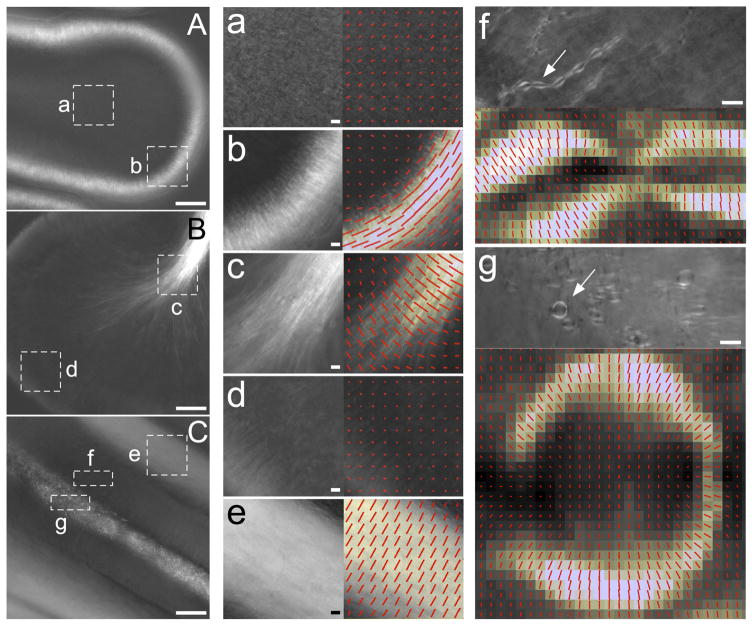
The polarized light microscope reveals orientational order in native molecular structures inside living cells, tissues, and whole organisms. It is a powerful tool used to monitor and analyze the early developmental stages of organisms that lend themselves to microscopic observations. In this article, we briefly discuss the components specific to a traditional polarizing microscope and some historically important observations on: chromosome packing in the sperm head, the first zygote division of the sea urchin, and differentiation initiated by the first asymmetric cell division in the sand dollar. We then introduce the LC-PolScope and describe its use for measuring birefringence and polarized fluorescence in living cells and tissues. Applications range from the enucleation of mouse oocytes to analyzing the polarized fluorescence of the water strider acrosome. We end with new results on the birefringence of the developing chick brain, which we analyzed between developmental stages of days 12-20.
© 2013 Wiley Periodicals, Inc.
Figures

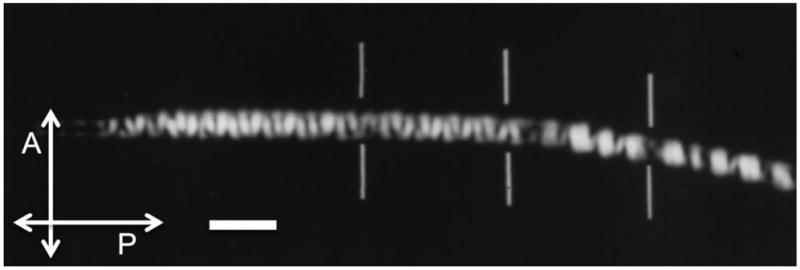




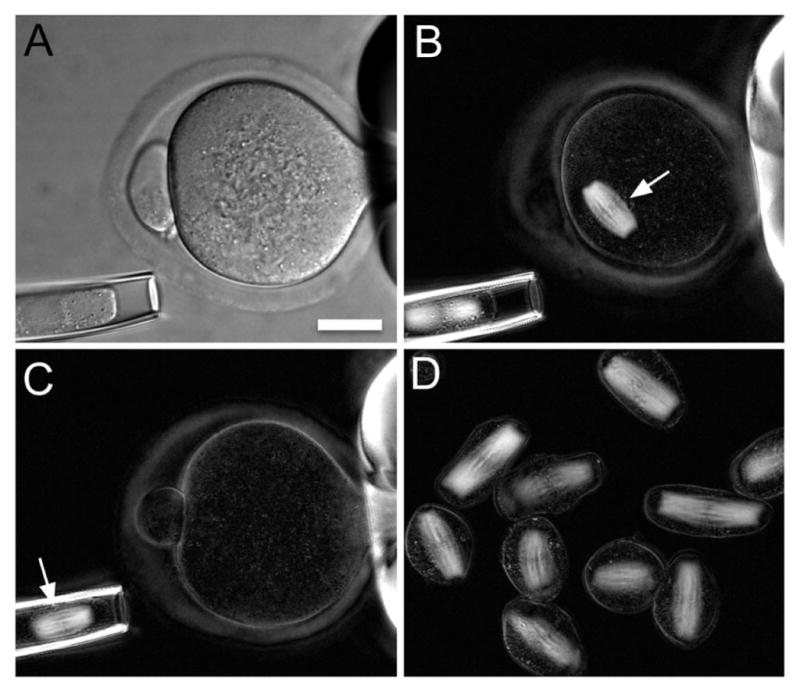
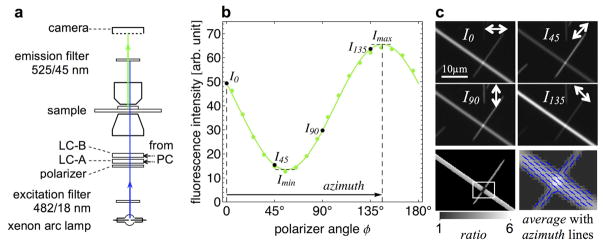
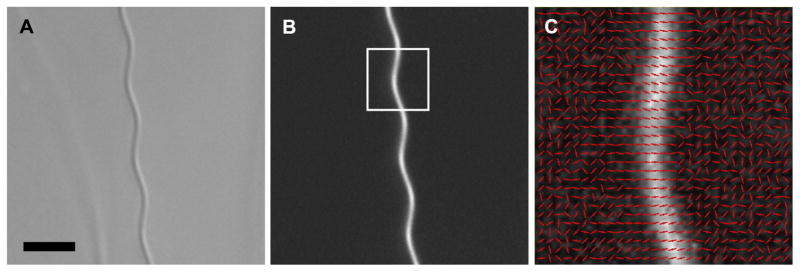


References
-
- Bennett HS. The microscopical investigation of biological materials with polarized light. In: Jones RM, editor. McClung’s handbook of microscopical techniques for workers in animal and plant tissues. 3. New York, NY: Paul B. Hoeber, Inc., Medical Book Department of Harper & Brothers; 1950. pp. 591–677.
-
- Bromage TG, Goldman HM, McFarlin SC, Warshaw J, Boyde A, Riggs CM. Circularly polarized light standards for investigations of collagen fiber orientation in bone. Anat Rec B New Anat. 2003;274:157–168. - PubMed
-
- Byrne JA, Pedersen DA, Clepper LL, Nelson M, Sanger WG, Gokhale S, Wolf DP, Mitalipov SM. Producing primate embryonic stem cells by somatic cell nuclear transfer. Nature. 2007;450:497–502. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
