

Completion of the entire hepatitis C virus life cycle in genetically humanized mice
- PMID: 23903655
- PMCID: PMC3858853
- DOI: 10.1038/nature12427
Completion of the entire hepatitis C virus life cycle in genetically humanized mice
Abstract
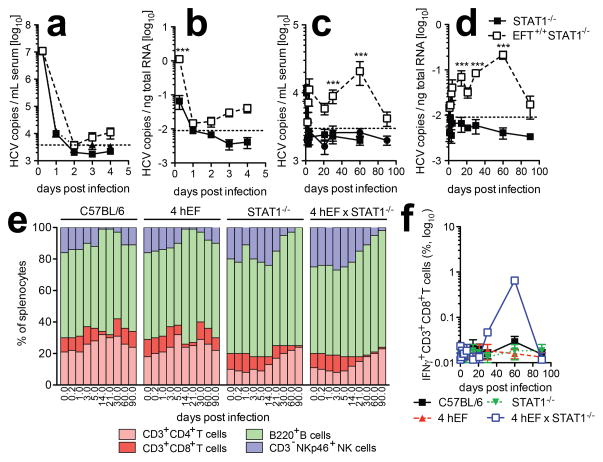
More than 130 million people worldwide chronically infected with hepatitis C virus (HCV) are at risk of developing severe liver disease. Antiviral treatments are only partially effective against HCV infection, and a vaccine is not available. Development of more efficient therapies has been hampered by the lack of a small animal model. Building on the observation that CD81 and occludin (OCLN) comprise the minimal set of human factors required to render mouse cells permissive to HCV entry, we previously showed that transient expression of these two human genes is sufficient to allow viral uptake into fully immunocompetent inbred mice. Here we demonstrate that transgenic mice stably expressing human CD81 and OCLN also support HCV entry, but innate and adaptive immune responses restrict HCV infection in vivo. Blunting antiviral immunity in genetically humanized mice infected with HCV results in measurable viraemia over several weeks. In mice lacking the essential cellular co-factor cyclophilin A (CypA), HCV RNA replication is markedly diminished, providing genetic evidence that this process is faithfully recapitulated. Using a cell-based fluorescent reporter activated by the NS3-4A protease we visualize HCV infection in single hepatocytes in vivo. Persistently infected mice produce de novo infectious particles, which can be inhibited with directly acting antiviral drug treatment, thereby providing evidence for the completion of the entire HCV life cycle in inbred mice. This genetically humanized mouse model opens new opportunities to dissect genetically HCV infection in vivo and provides an important preclinical platform for testing and prioritizing drug candidates and may also have utility for evaluating vaccine efficacy.
Conflict of interest statement
Figures




Comment in
-
Liver disease: Quest to find new mouse models for liver diseases.Nat Rev Gastroenterol Hepatol. 2013 Sep;10(9):501. doi: 10.1038/nrgastro.2013.153. Epub 2013 Aug 13. Nat Rev Gastroenterol Hepatol. 2013. PMID: 23938451 No abstract available.
-
Genetically humanized mice recapitulate the entire hepatitis C virus life cycle.J Hepatol. 2014 Mar;60(3):671-3. doi: 10.1016/j.jhep.2013.10.005. Epub 2013 Oct 12. J Hepatol. 2014. PMID: 24128418 No abstract available.
-
Establishment of mice with inheritable susceptibility to productive hepatitis C virus infection.Hepatology. 2014 May;59(5):2043-6. doi: 10.1002/hep.26949. Epub 2014 Mar 27. Hepatology. 2014. PMID: 24277664 No abstract available.
References
-
- Barth H, et al. Cellular binding of hepatitis C virus envelope glycoprotein E2 requires cell surface heparan sulfate. J Biol Chem. 2003;278:41003–41012. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
