

Nonuniform chain collapse during early stages of staphylococcal nuclease folding detected by fluorescence resonance energy transfer and ultrarapid mixing methods
- PMID: 23904284
- PMCID: PMC3795492
- DOI: 10.1002/pro.2320
Nonuniform chain collapse during early stages of staphylococcal nuclease folding detected by fluorescence resonance energy transfer and ultrarapid mixing methods
Abstract
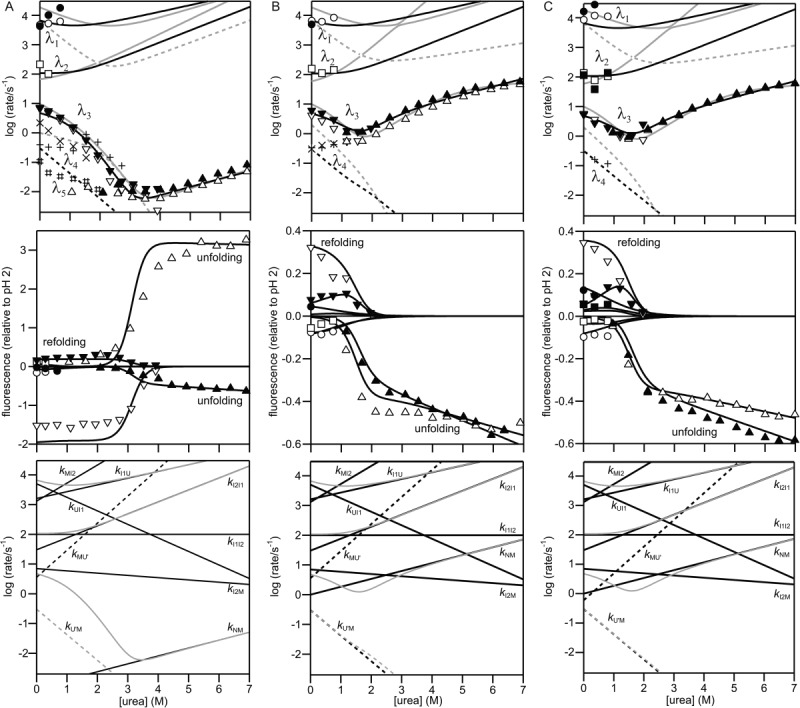
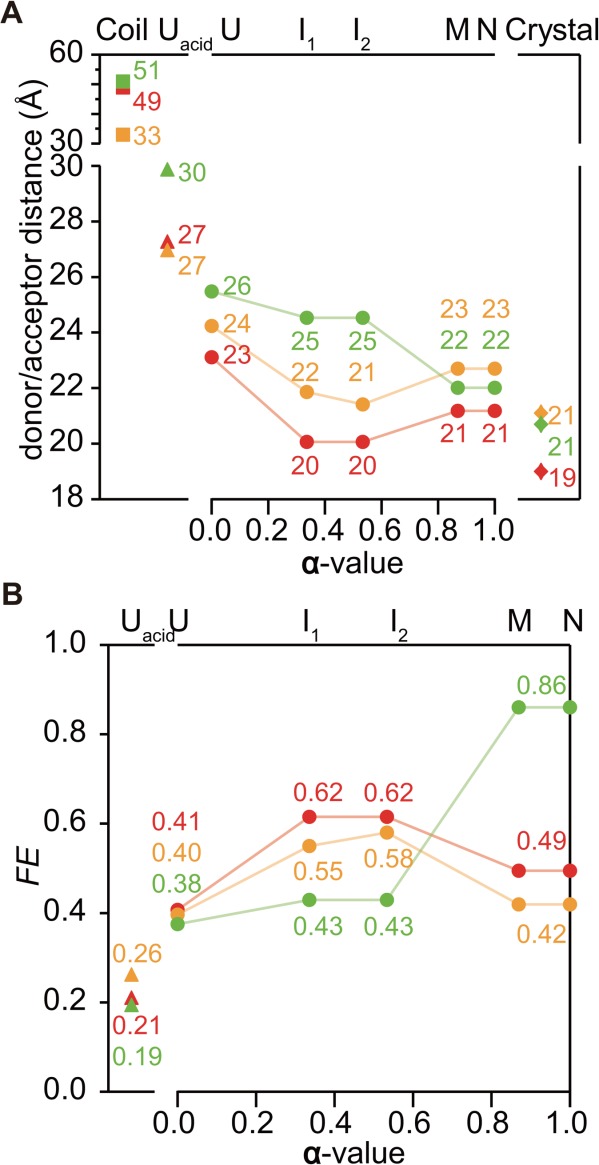
The development of tertiary structure during folding of staphylococcal nuclease (SNase) was studied by time-resolved fluorescence resonance energy transfer measured using continuous- and stopped-flow techniques. Variants of this two-domain protein containing intradomain and interdomain fluorescence donor/acceptor pairs (Trp and Cys-linked fluorophore or quencher) were prepared to probe the intradomain and interdomain structural evolution accompanying SNase folding. The intra-domain donor/acceptor pairs are within the β-barrel domain (Trp27/Cys64 and Trp27/Cys97) and the interdomain pair is between the α-helical domain and the β-barrel domain (Trp140/Cys64). Time-resolved energy transfer efficiency accompanying folding and unfolding at different urea concentrations was measured over a time range from 30 μs to ≈ 10 s. Information on average donor/acceptor distances at different stages of the folding process was obtained by using a quantitative kinetic modeling approach. The average distance for the donor/acceptor pairs in the β-barrel domain decreases to nearly native values whereas that of the interdomain donor/acceptor pairs remains unchanged in the earliest intermediate (<500 μs of refolding). This indicates a rapid nonuniform collapse resulting in an ensemble of heterogeneous conformations in which the central region of the β-barrel domain is well developed while the C-terminal α-helical domain remains disordered. The distance between Trp140 and Cys64 decreases to native values on the 100-ms time scale, indicating that the α-helical domain docks onto the preformed β-barrel at a late stage of the folding. In addition, the unfolded state is found to be more compact under native conditions, suggesting that changes in solvent conditions may induce a nonspecific hydrophobic collapse.
Keywords: Förster energy transfer; continuous-flow; fluorescence; protein folding; stopped-flow.
© 2013 The Protein Society.
Figures





References
-
- Baldwin RL, Rose GD. Is protein folding hierarchic? II. Folding intermediates and transition states. Trends Biochem Sci. 1999;24:77–83. - PubMed
-
- Arai M, Kuwajima K. Role of the molten globule state in protein folding. Adv Protein Chem. 2000;53:209–282. - PubMed
-
- Udgaonkar JB, Baldwin RL. NMR evidence for an early framework intermediate on the folding pathway of ribonuclease A. Nature. 1988;335:694–699. - PubMed
-
- Roder H, Wüthrich K. Protein folding kinetics by combined use of rapid mixing techniques and NMR observation of individual amide protons. Proteins. 1986;1:34–42. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
