

Functional lysine modification by an intrinsically reactive primary glycolytic metabolite
- PMID: 23908237
- PMCID: PMC4005992
- DOI: 10.1126/science.1238327
Functional lysine modification by an intrinsically reactive primary glycolytic metabolite
Abstract
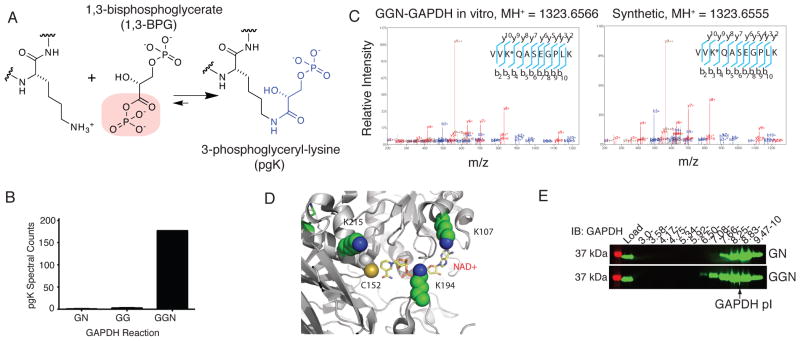
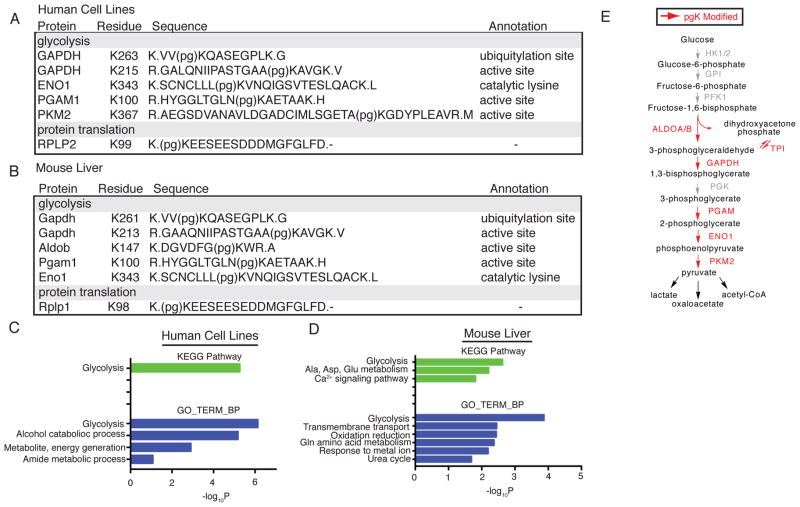
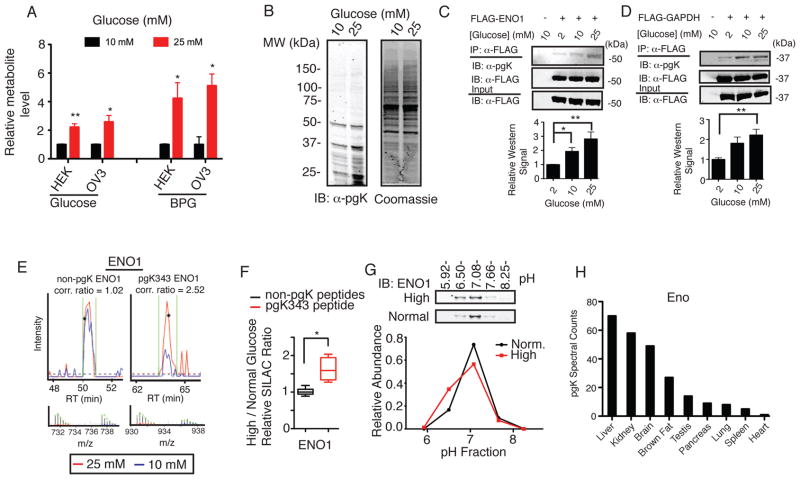
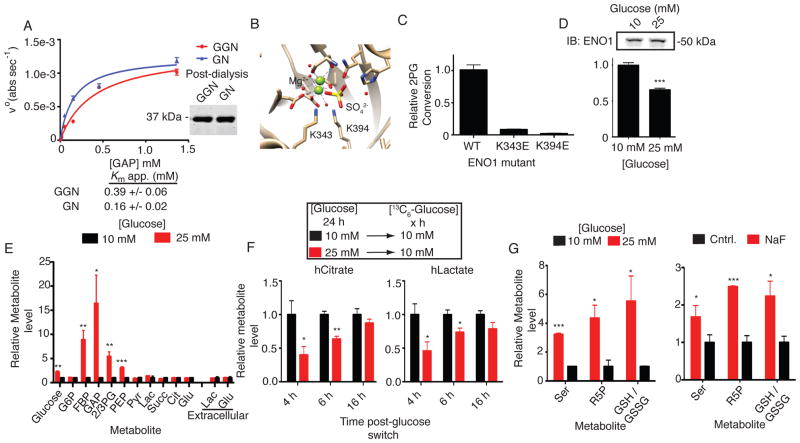
The posttranslational modification of proteins and their regulation by metabolites represent conserved mechanisms in biology. At the confluence of these two processes, we report that the primary glycolytic intermediate 1,3-bisphosphoglycerate (1,3-BPG) reacts with select lysine residues in proteins to form 3-phosphoglyceryl-lysine (pgK). This reaction, which does not require enzyme catalysis, but rather exploits the electrophilicity of 1,3-BPG, was found by proteomic profiling to be enriched on diverse classes of proteins and prominently in or around the active sites of glycolytic enzymes. pgK modifications inhibit glycolytic enzymes and, in cells exposed to high glucose, accumulate on these enzymes to create a potential feedback mechanism that contributes to the buildup and redirection of glycolytic intermediates to alternate biosynthetic pathways.
Figures
References
-
- Sola-Penna M, Da Silva D, Coelho WS, Marinho-Carvalho MM, Zancan P. Regulation of mammalian muscle type 6-phosphofructo-1-kinase and its implication for the control of the metabolism. IUBMB life. 2010 Nov;62:791. - PubMed
-
- Walsh C. Posttranslational modification of proteins : expanding nature’s inventory. Roberts and Co. Publishers; Englewood, Colo: 2006. p. xxi.p. 490.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
