

Proximal tubule function and response to acidosis
- PMID: 23908456
- PMCID: PMC4152816
- DOI: 10.2215/CJN.10391012
Proximal tubule function and response to acidosis
Abstract
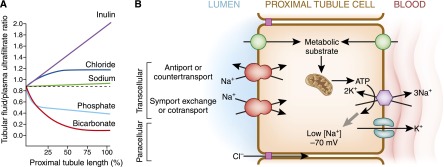
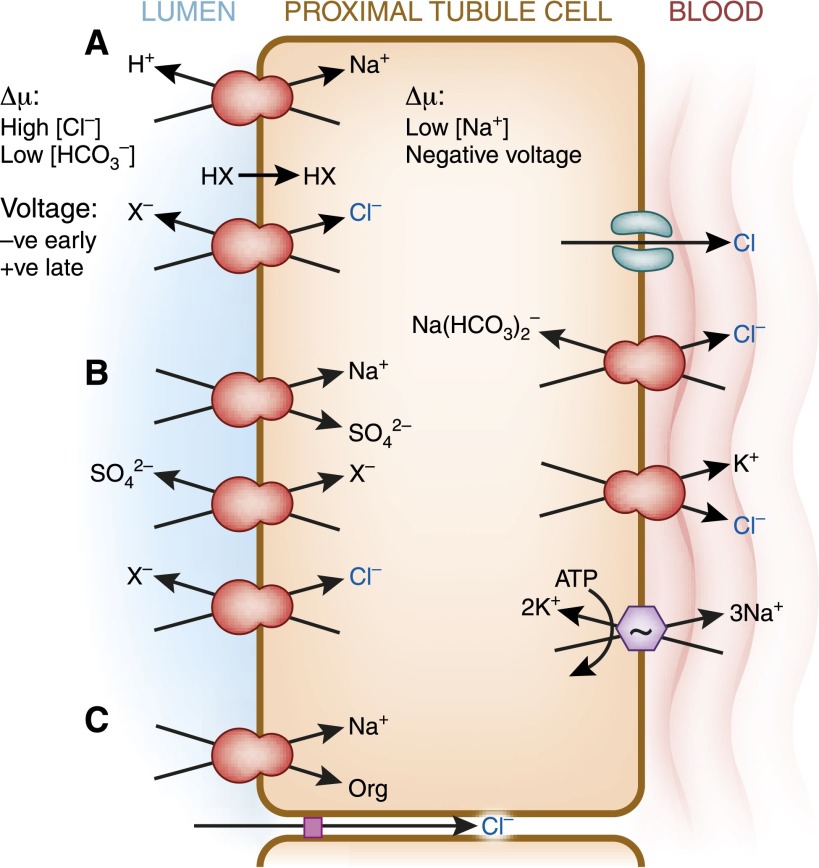
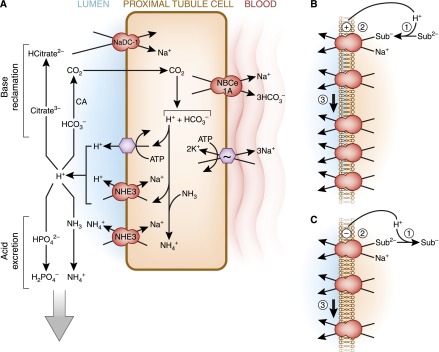
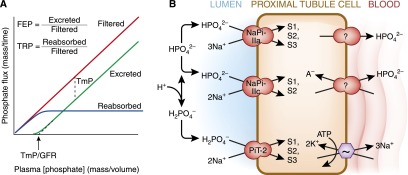
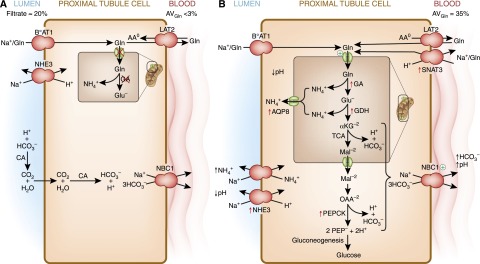
The human kidneys produce approximately 160-170 L of ultrafiltrate per day. The proximal tubule contributes to fluid, electrolyte, and nutrient homeostasis by reabsorbing approximately 60%-70% of the water and NaCl, a greater proportion of the NaHCO3, and nearly all of the nutrients in the ultrafiltrate. The proximal tubule is also the site of active solute secretion, hormone production, and many of the metabolic functions of the kidney. This review discusses the transport of NaCl, NaHCO3, glucose, amino acids, and two clinically important anions, citrate and phosphate. NaCl and the accompanying water are reabsorbed in an isotonic fashion. The energy that drives this process is generated largely by the basolateral Na(+)/K(+)-ATPase, which creates an inward negative membrane potential and Na(+)-gradient. Various Na(+)-dependent countertransporters and cotransporters use the energy of this gradient to promote the uptake of HCO3 (-) and various solutes, respectively. A Na(+)-dependent cotransporter mediates the movement of HCO3 (-) across the basolateral membrane, whereas various Na(+)-independent passive transporters accomplish the export of various other solutes. To illustrate its homeostatic feat, the proximal tubule alters its metabolism and transport properties in response to metabolic acidosis. The uptake and catabolism of glutamine and citrate are increased during acidosis, whereas the recovery of phosphate from the ultrafiltrate is decreased. The increased catabolism of glutamine results in increased ammoniagenesis and gluconeogenesis. Excretion of the resulting ammonium ions facilitates the excretion of acid, whereas the combined pathways accomplish the net production of HCO3 (-) ions that are added to the plasma to partially restore acid-base balance.
Copyright © 2014 by the American Society of Nephrology.
Figures
References
-
- Jones G, Prosser DE, Kaufmann M: 25-Hydroxyvitamin D-24-hydroxylase (CYP24A1): Its important role in the degradation of vitamin D. Arch Biochem Biophys 523: 9–18, 2012 - PubMed
-
- Meyer C, Dostou JM, Gerich JE: Role of the human kidney in glucose counterregulation. Diabetes 48: 943–948, 1999 - PubMed
-
- De Vito E, Cabrera RR, Fasciolo JC: Renin production and release by rat kidney slices. Am J Physiol 219: 1042–1045, 1970 - PubMed
-
- Koury ST, Bondurant MC, Koury MJ: Localization of erythropoietin synthesizing cells in murine kidneys by in situ hybridization. Blood 71: 524–527, 1988 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
