

TASK-2: a K2P K(+) channel with complex regulation and diverse physiological functions
- PMID: 23908634
- PMCID: PMC3725403
- DOI: 10.3389/fphys.2013.00198
TASK-2: a K2P K(+) channel with complex regulation and diverse physiological functions
Abstract
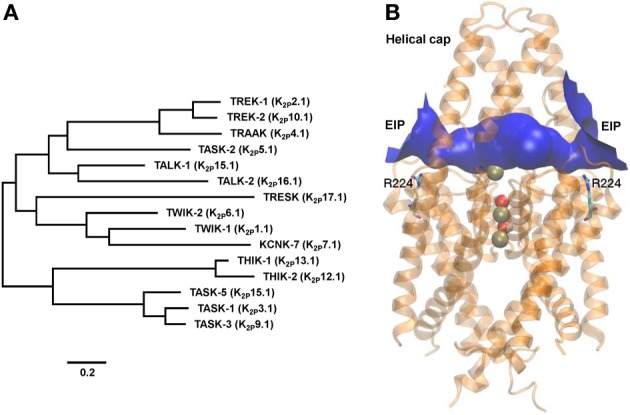
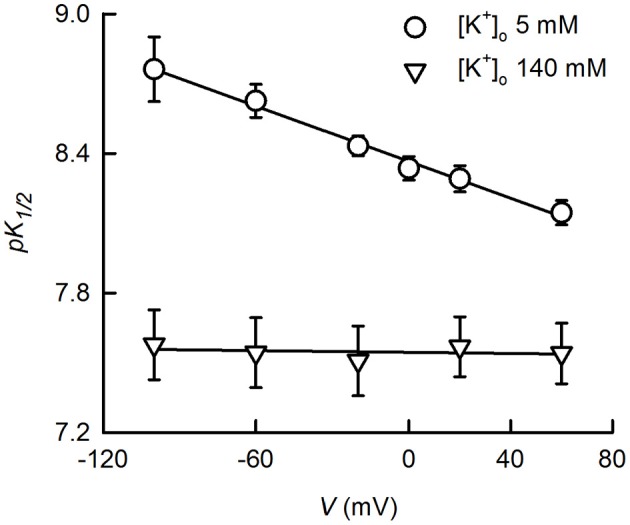
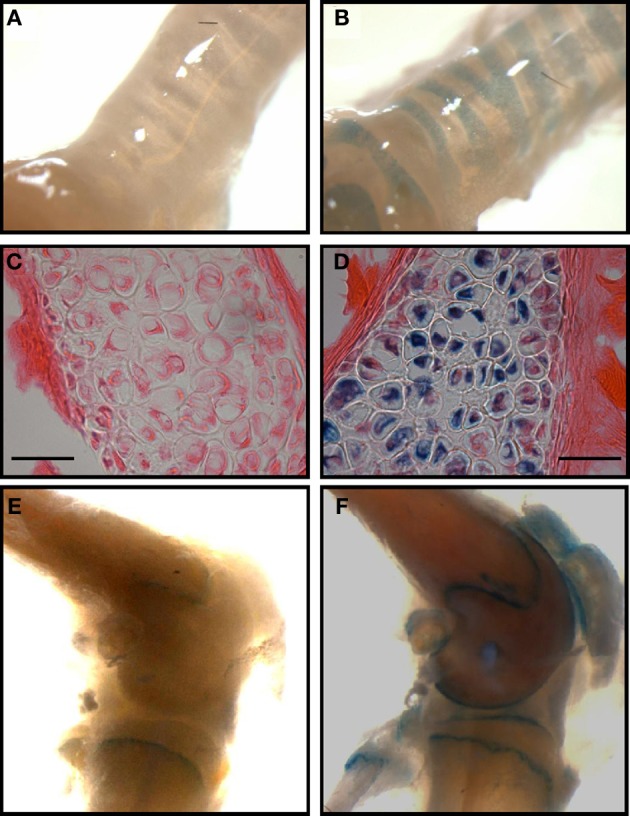
TASK-2 (K2P5.1) is a two-pore domain K(+) channel belonging to the TALK subgroup of the K2P family of proteins. TASK-2 has been shown to be activated by extra- and intracellular alkalinization. Extra- and intracellular pH-sensors reside at arginine 224 and lysine 245 and might affect separate selectivity filter and inner gates respectively. TASK-2 is modulated by changes in cell volume and a regulation by direct G-protein interaction has also been proposed. Activation by extracellular alkalinization has been associated with a role of TASK-2 in kidney proximal tubule bicarbonate reabsorption, whilst intracellular pH-sensitivity might be the mechanism for its participation in central chemosensitive neurons. In addition to these functions TASK-2 has been proposed to play a part in apoptotic volume decrease in kidney cells and in volume regulation of glial cells and T-lymphocytes. TASK-2 is present in chondrocytes of hyaline cartilage, where it is proposed to play a central role in stabilizing the membrane potential. Additional sites of expression are dorsal root ganglion neurons, endocrine and exocrine pancreas and intestinal smooth muscle cells. TASK-2 has been associated with the regulation of proliferation of breast cancer cells and could become target for breast cancer therapeutics. Further work in native tissues and cells together with genetic modification will no doubt reveal the details of TASK-2 functions that we are only starting to suspect.
Keywords: K2P channels; TASK-2 channel; bicarbonate reabsorption; cell volume regulation; central chemoception; chondrocytes.
Figures

References
-
- Alper S. L. (2010). Familial renal tubular acidosis. J. Nephrol. 23Suppl. 16, S57–S76 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources