

Analysis, Isolation, and Activation of Antigen-Specific CD4(+) and CD8(+) T Cells by Soluble MHC-Peptide Complexes
- PMID: 23908656
- PMCID: PMC3726995
- DOI: 10.3389/fimmu.2013.00218
Analysis, Isolation, and Activation of Antigen-Specific CD4(+) and CD8(+) T Cells by Soluble MHC-Peptide Complexes
Abstract
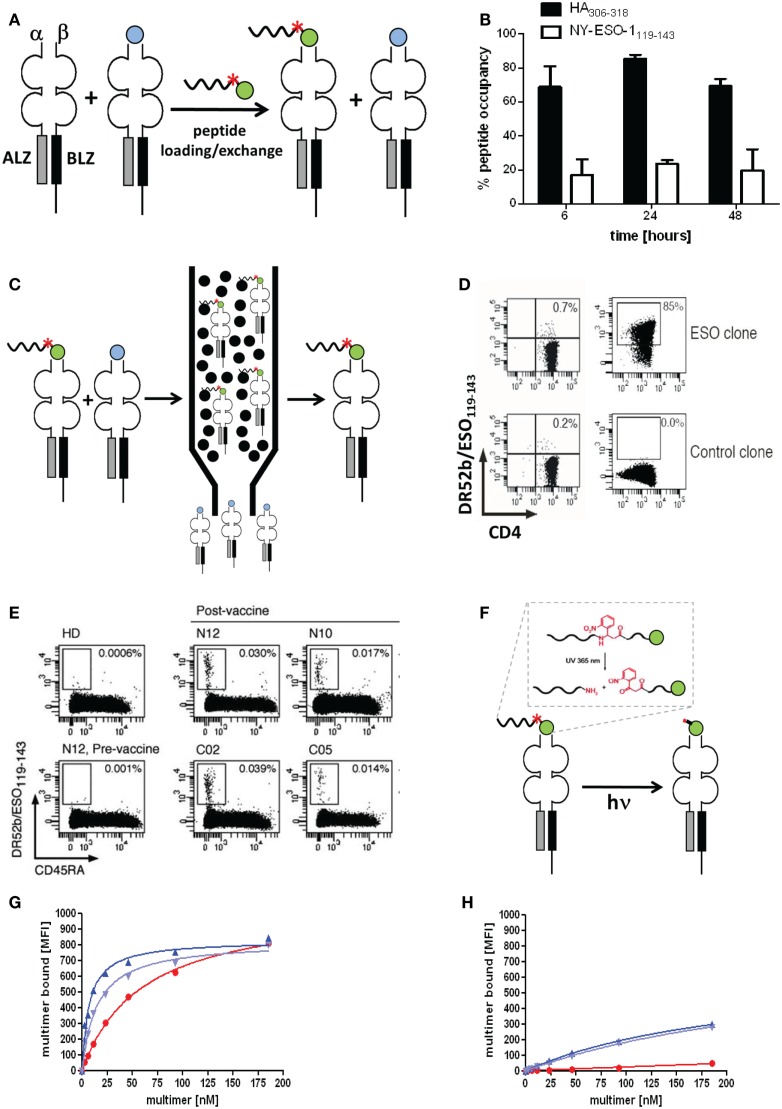
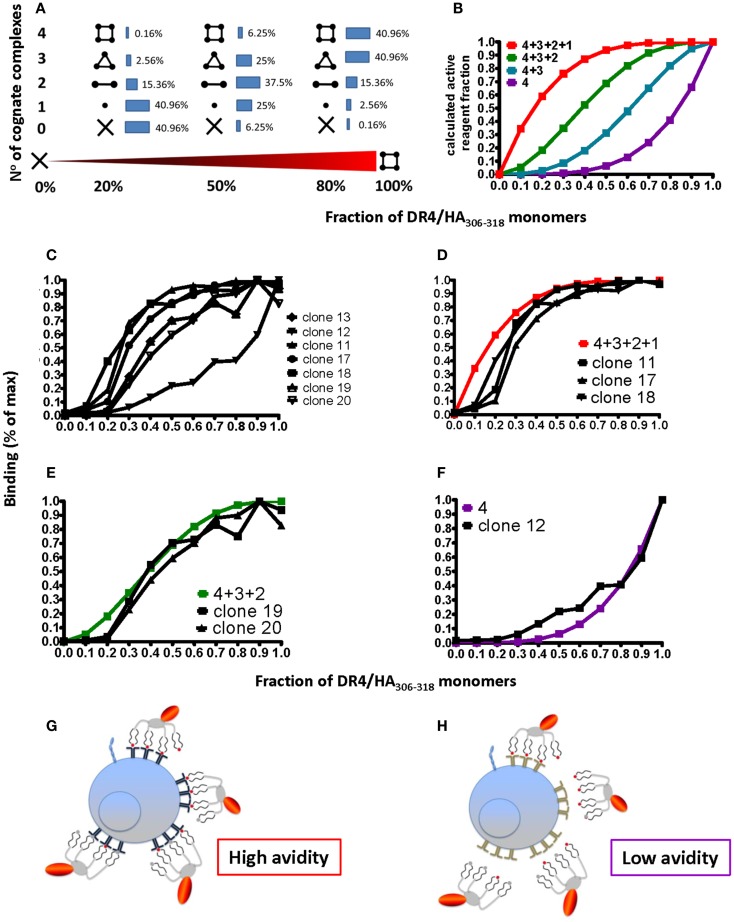
T cells constitute the core of adaptive cellular immunity and protect higher organisms against pathogen infections and cancer. Monitoring of disease progression as well as prophylactic or therapeutic vaccines and immunotherapies call for conclusive detection, analysis, and sorting of antigen-specific T cells. This is possible by means of soluble recombinant ligands for T cells, i.e., MHC class I-peptide (pMHC I) complexes for CD8(+) T cells and MHC class II-peptide (pMHC II) complexes for CD4(+) T cells and flow cytometry. Here we review major developments in the development of pMHC staining reagents and their diverse applications and discuss perspectives of their use for basic and clinical investigations.
Keywords: MHC; T cell receptor; T cells; coreceptor; flow cytometry; tetramers.
Figures
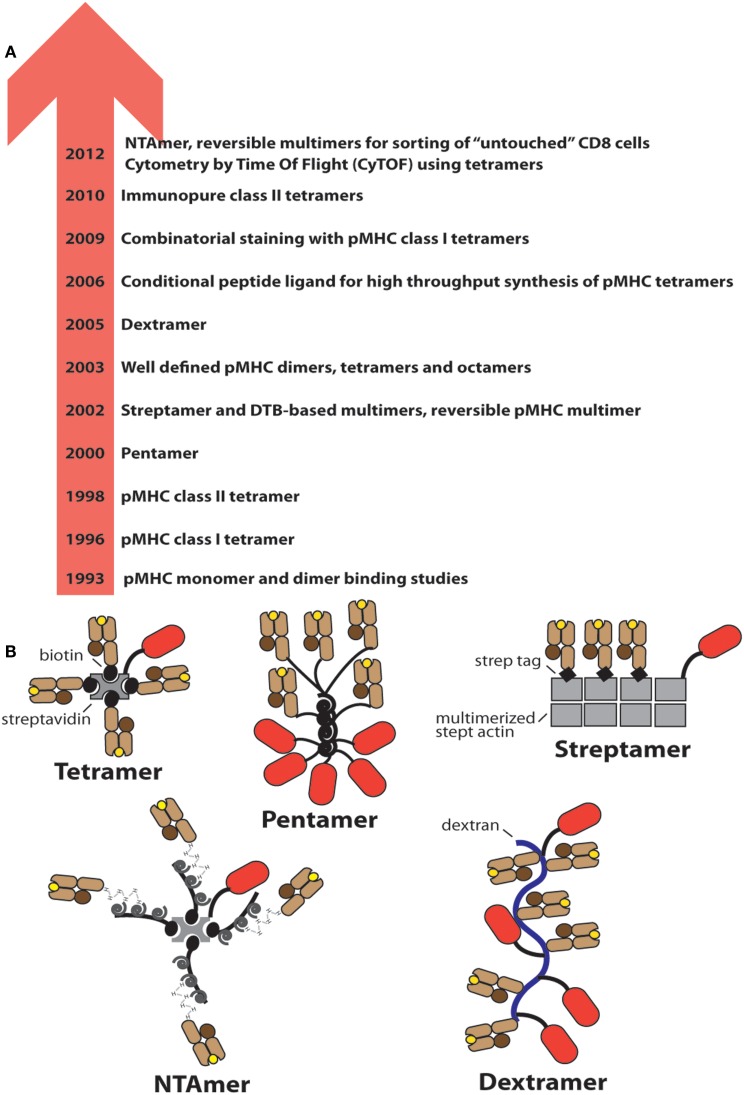
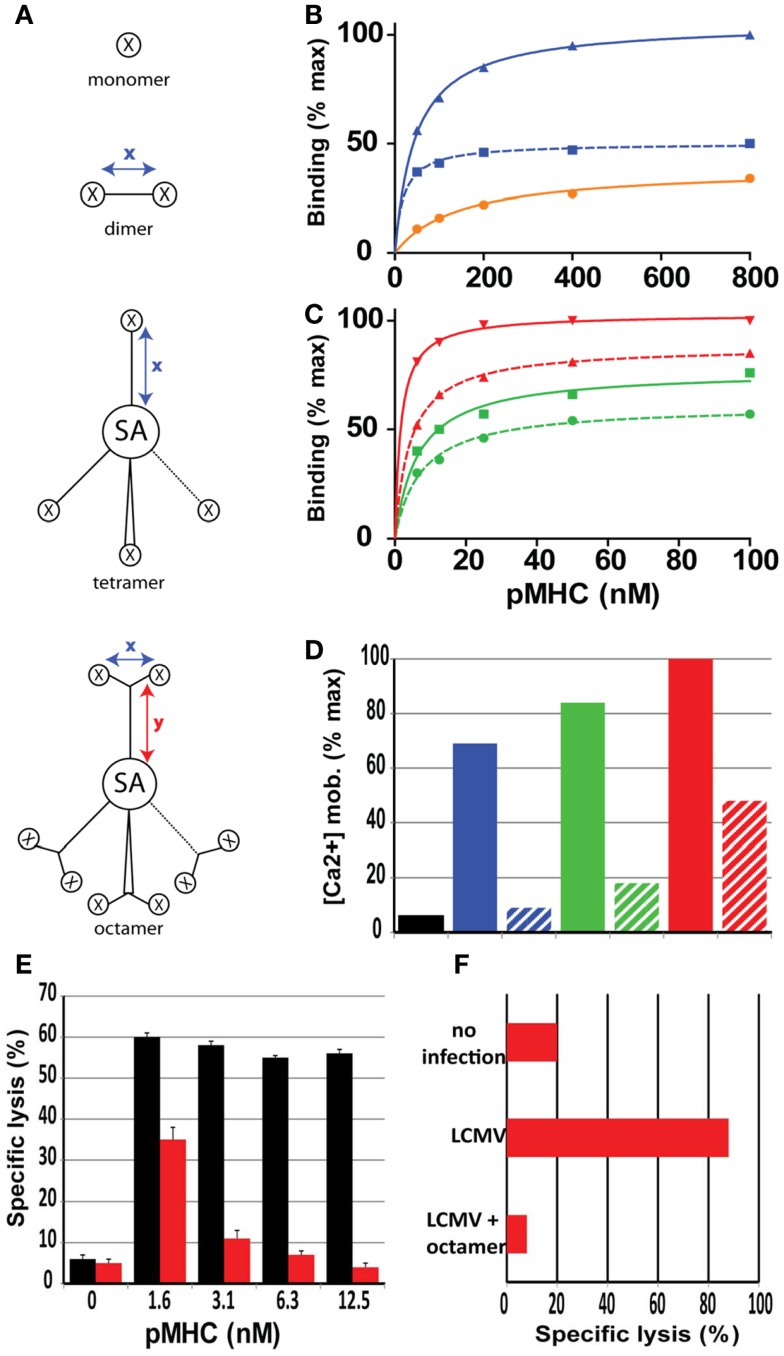
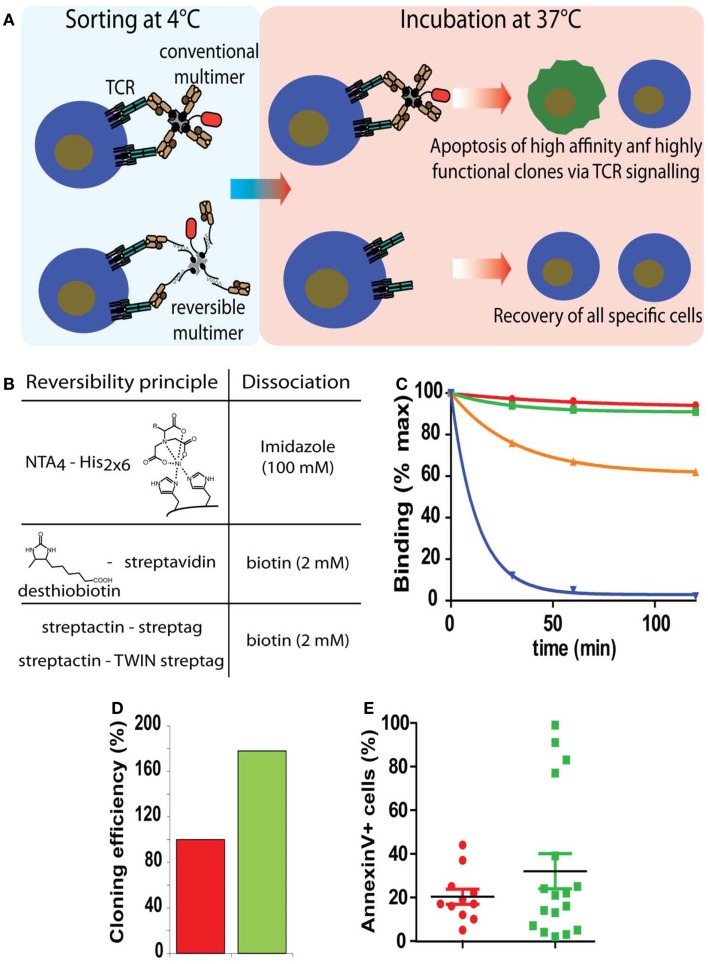
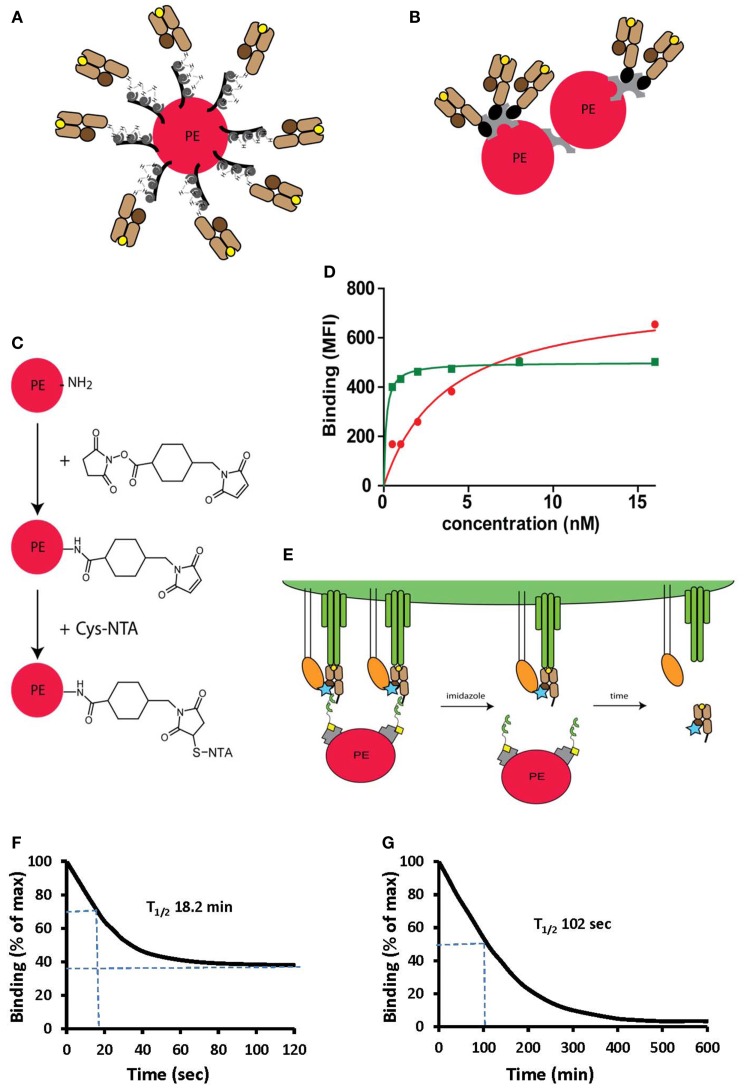
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous