

Functional genomic characterization of neoblast-like stem cells in larval Schistosoma mansoni
- PMID: 23908765
- PMCID: PMC3728622
- DOI: 10.7554/eLife.00768
Functional genomic characterization of neoblast-like stem cells in larval Schistosoma mansoni
Abstract
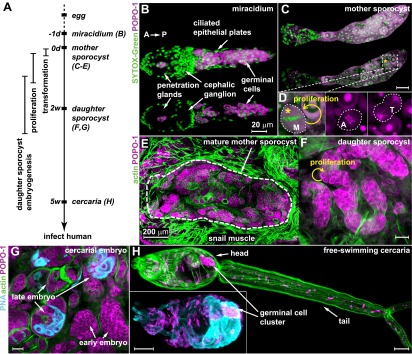
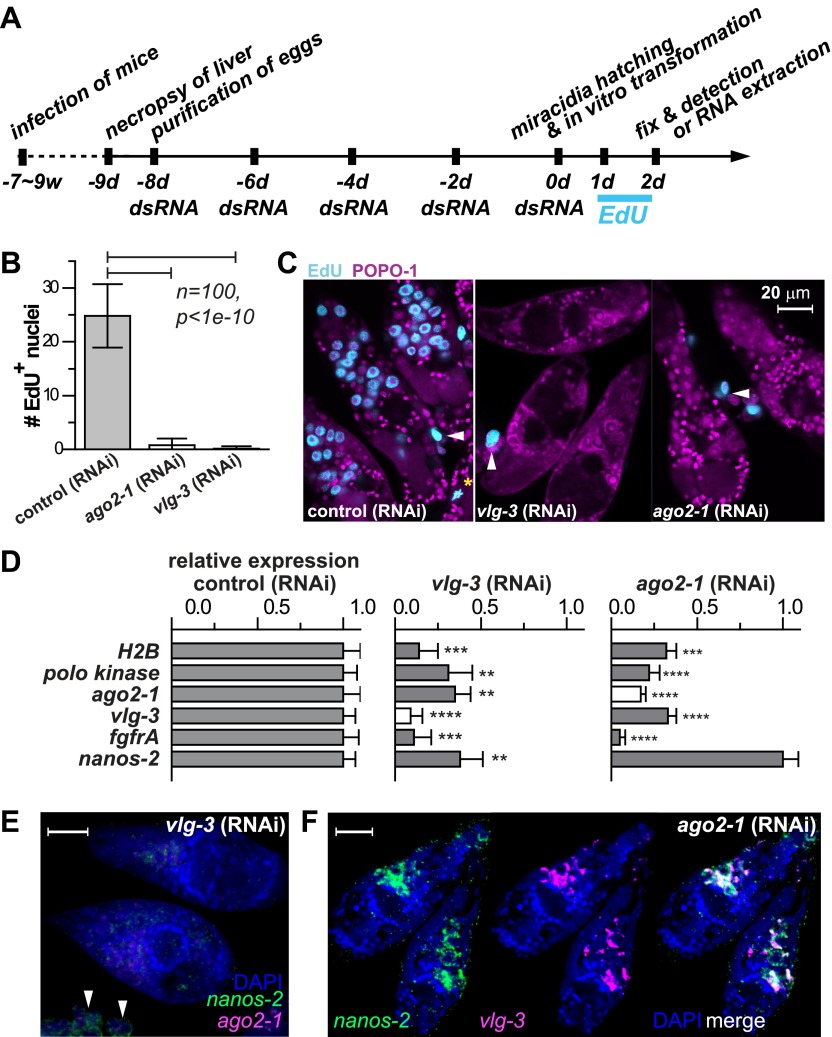
Schistosomes infect hundreds of millions of people in the developing world. Transmission of these parasites relies on a stem cell-driven, clonal expansion of larvae inside a molluscan intermediate host. How this novel asexual reproductive strategy relates to current models of stem cell maintenance and germline specification is unclear. Here, we demonstrate that this proliferative larval cell population (germinal cells) shares some molecular signatures with stem cells from diverse organisms, in particular neoblasts of planarians (free-living relatives of schistosomes). We identify two distinct germinal cell lineages that differ in their proliferation kinetics and expression of a nanos ortholog. We show that a vasa/PL10 homolog is required for proliferation and maintenance of both populations, whereas argonaute2 and a fibroblast growth factor receptor-encoding gene are required only for nanos-negative cells. Our results suggest that an ancient stem cell-based developmental program may have enabled the evolution of the complex life cycle of parasitic flatworms. DOI:http://dx.doi.org/10.7554/eLife.00768.001.
Keywords: Other; Schistosoma mansoni; neoblasts; schistosomiasis; stem cells; trematode development.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures






Comment in
-
On the trail of a tropical disease.Elife. 2013 Jul 30;2:e01115. doi: 10.7554/eLife.01115. Elife. 2013. PMID: 23908770 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
