

Mathematical modeling of eukaryotic cell migration: insights beyond experiments
- PMID: 23909278
- PMCID: PMC4148455
- DOI: 10.1146/annurev-cellbio-101512-122308
Mathematical modeling of eukaryotic cell migration: insights beyond experiments
Abstract
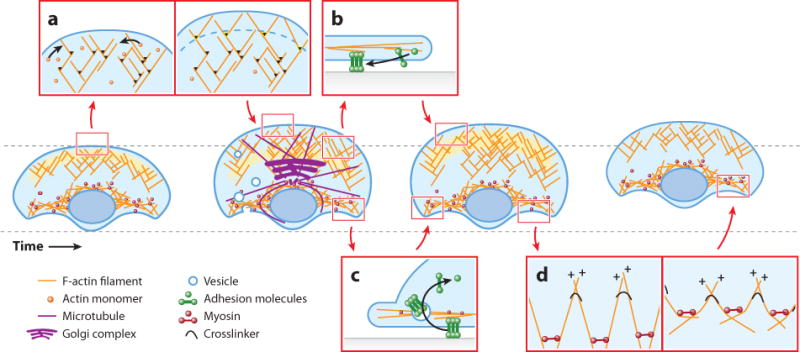
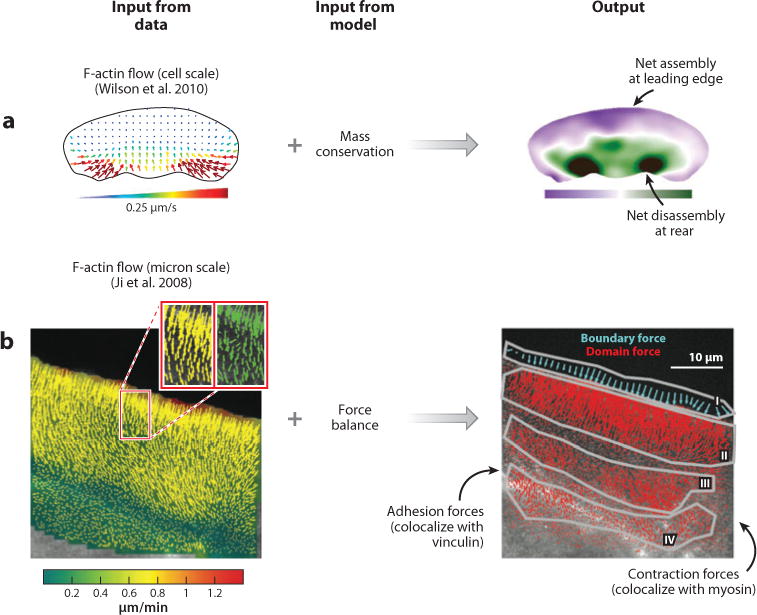
A migrating cell is a molecular machine made of tens of thousands of short-lived and interacting parts. Understanding migration means understanding the self-organization of these parts into a system of functional units. This task is one of tackling complexity: First, the system integrates numerous chemical and mechanical component processes. Second, these processes are connected in feedback interactions and over a large range of spatial and temporal scales. Third, many processes are stochastic, which leads to heterogeneous migration behaviors. Early on in the research of cell migration it became evident that this complexity exceeds human intuition. Thus, the cell migration community has led the charge to build mathematical models that could integrate the diverse experimental observations and measurements in consistent frameworks, first in conceptual and more recently in molecularly explicit models. The main goal of this review is to sift through a series of important conceptual and explicit mathematical models of cell migration and to evaluate their contribution to the field in their ability to integrate critical experimental data.
Figures



References
-
- Abercrombie M. The Croonian lecture, 1978: the crawling movement of metazoan cells. Proc R Soc Lond B. 1980;207:129–47.
-
- Allena R. Cell migration with multiple pseudopodia: temporal and spatial sensing models. Bull Math Biol. 2013;75:288–316. - PubMed
-
- Asnacios A, Hamant O. The mechanics behind cell polarity. Trends Cell Biol. 2012;22:584–91. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
