

Seeing the unseen: the hidden world of slow axonal transport
- PMID: 23912032
- PMCID: PMC3902140
- DOI: 10.1177/1073858413498306
Seeing the unseen: the hidden world of slow axonal transport
Abstract
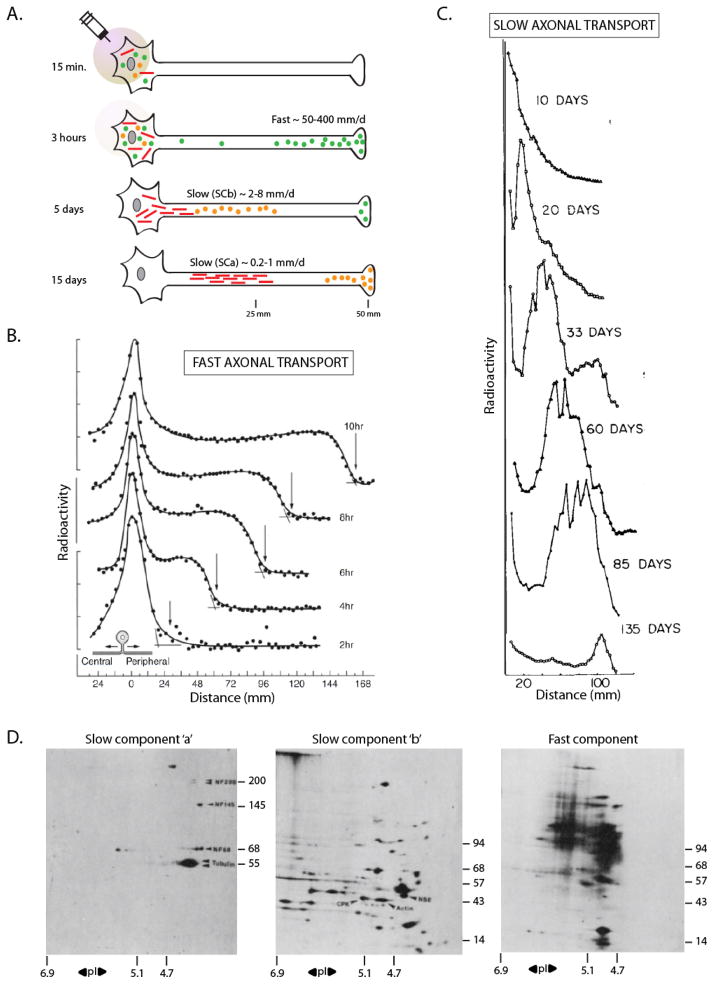
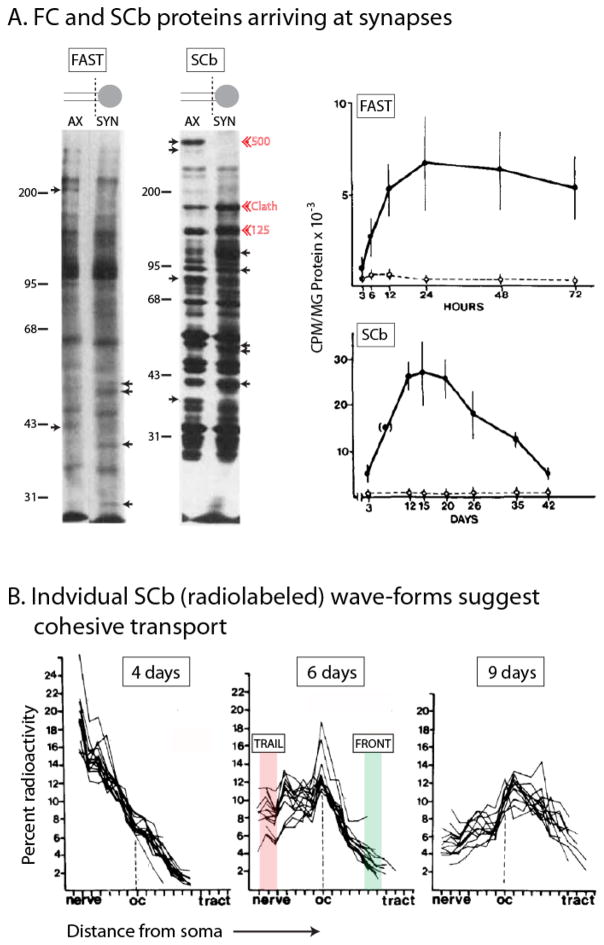
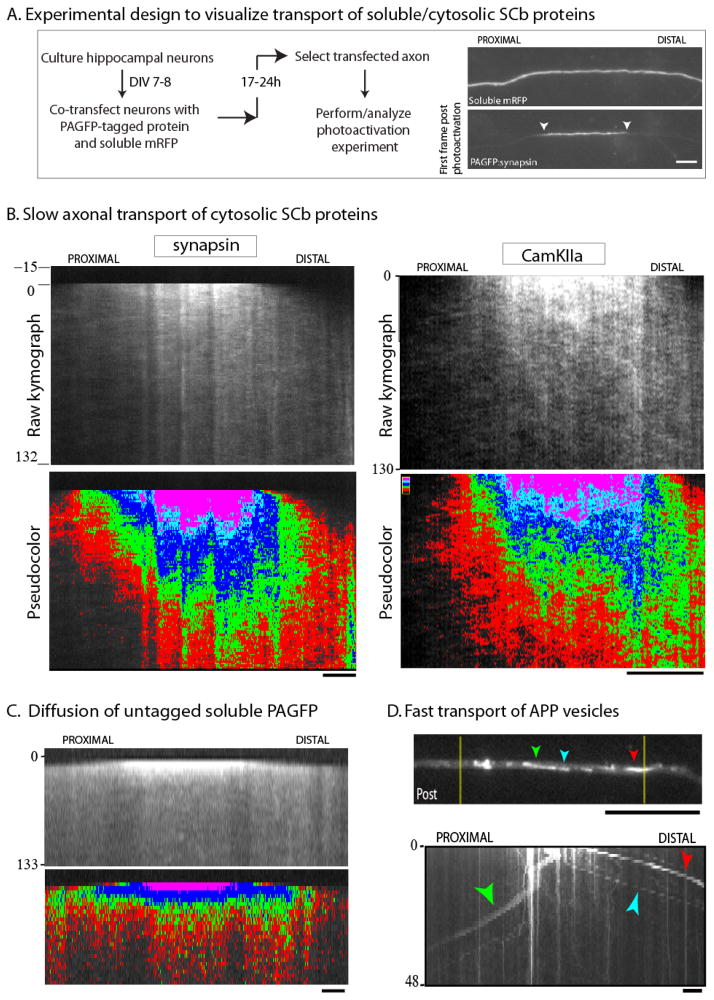
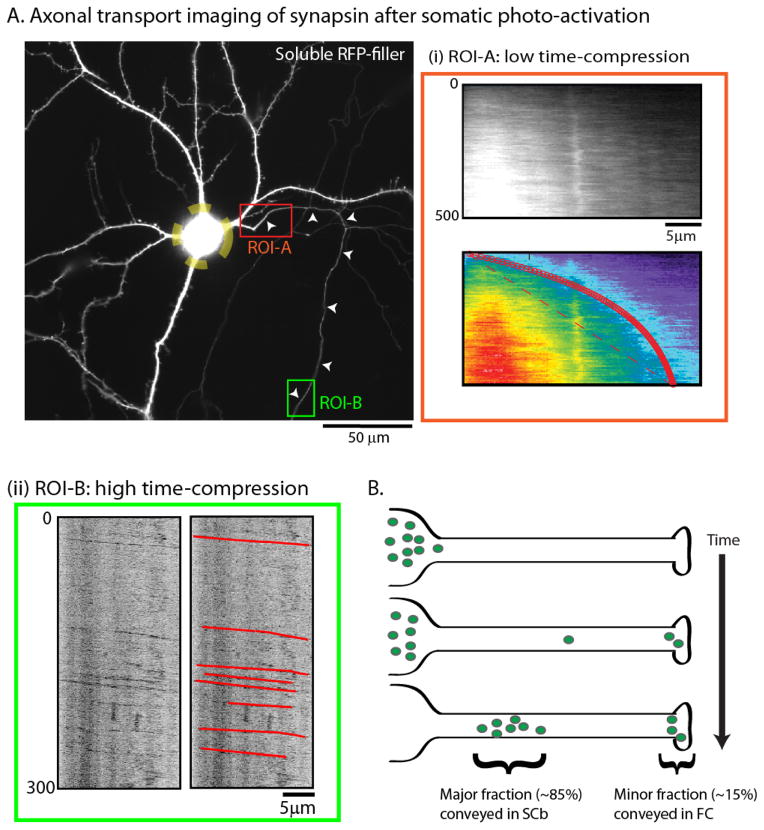
Axonal transport is the lifeline of axons and synapses. After synthesis in neuronal cell bodies, proteins are conveyed into axons in two distinct rate classes-fast and slow axonal transport. Whereas fast transport delivers vesicular cargoes, slow transport carries cytoskeletal and cytosolic (or soluble) proteins that have critical roles in neuronal structure and function. Although significant progress has been made in dissecting the molecular mechanisms of fast vesicle transport, mechanisms of slow axonal transport are less clear. Why is this so? Historically, conceptual advances in the axonal transport field have paralleled innovations in imaging the movement, and slow-transport cargoes are not as readily seen as motile vesicles. However, new ways of visualizing slow transport have reenergized the field, leading to fundamental insights that have changed our views on axonal transport, motor regulation, and intracellular trafficking in general. This review first summarizes classic studies that characterized axonal transport, and then discusses recent technical and conceptual advances in slow axonal transport that have provided insights into some long-standing mysteries.
Keywords: cargo complexes; cytosolic proteins; diffusion; slow axonal transport; soluble proteins; transport packets.
Conflict of interest statement
Conflict of interest: none
Figures
References
-
- Allen RD, Metuzals J, Tasaki I, Brady ST, Gilbert SP. Fast axonal transport in squid giant axon. Science. 1982;218:1127–1129. - PubMed
-
- Baas PW, Brown A. Slow axonal transport: the polymer transport model. Trends Cell Biol. 1997;7:380–384. - PubMed
-
- Black MM, Lasek RJ. Axonal transport of actin: slow component b is the principal source of actin for the axon. Brain Res. 1979;171:401–13. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
