

Inhibition of lipopolysaccharide-induced proinflammatory responses by Buddleja officinalis extract in BV-2 microglial cells via negative regulation of NF-kB and ERK1/2 signaling
- PMID: 23912273
- PMCID: PMC6270610
- DOI: 10.3390/molecules18089195
Inhibition of lipopolysaccharide-induced proinflammatory responses by Buddleja officinalis extract in BV-2 microglial cells via negative regulation of NF-kB and ERK1/2 signaling
Abstract
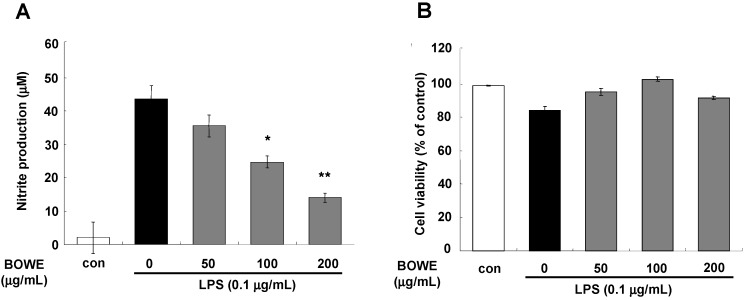
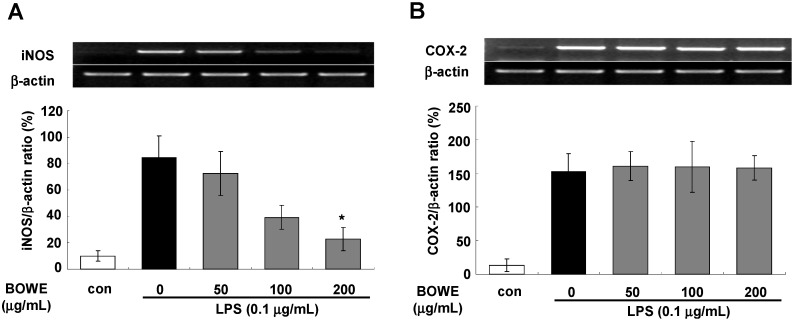
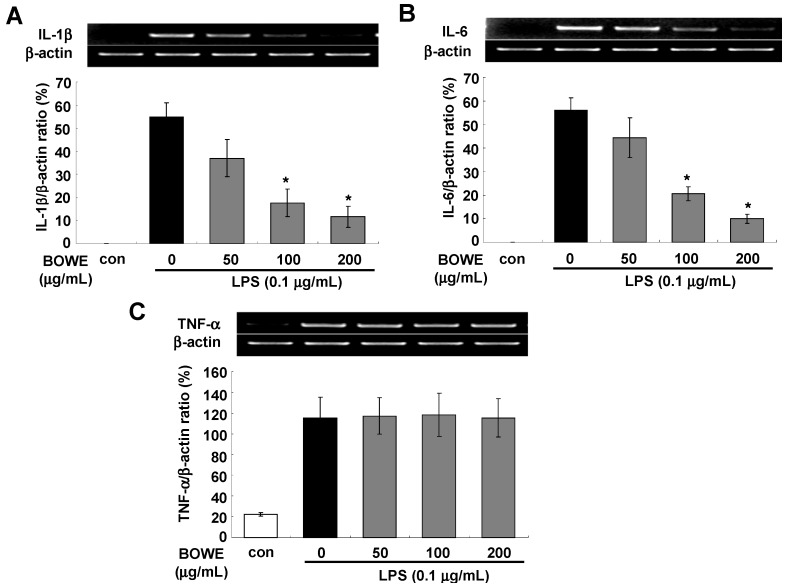
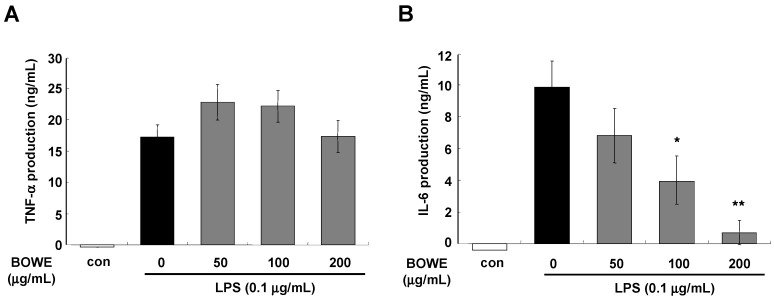
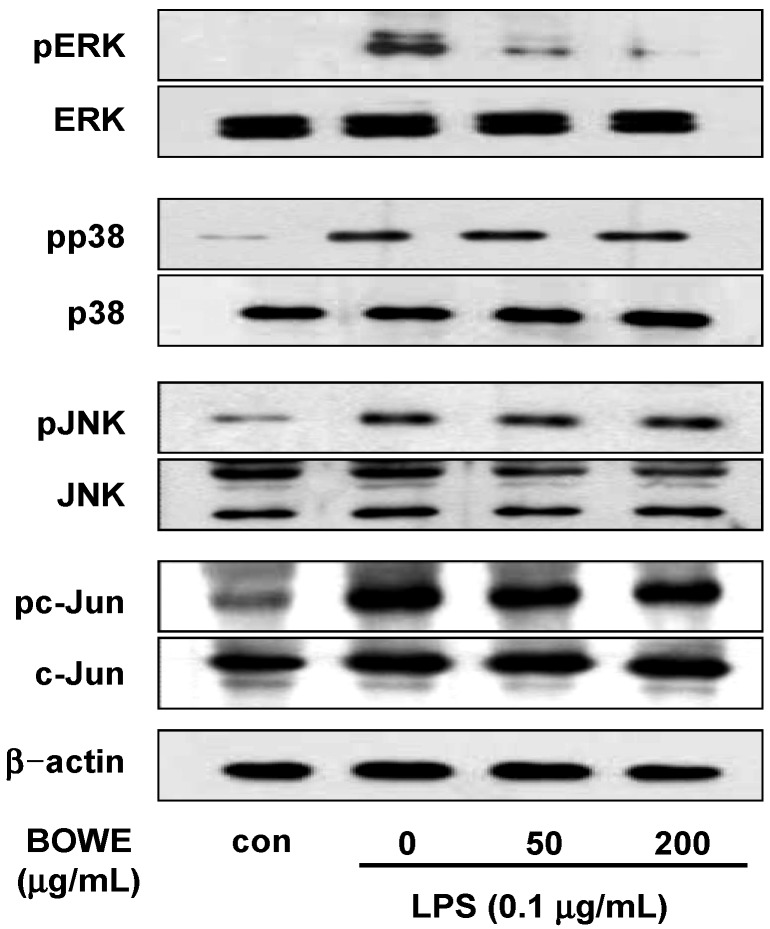
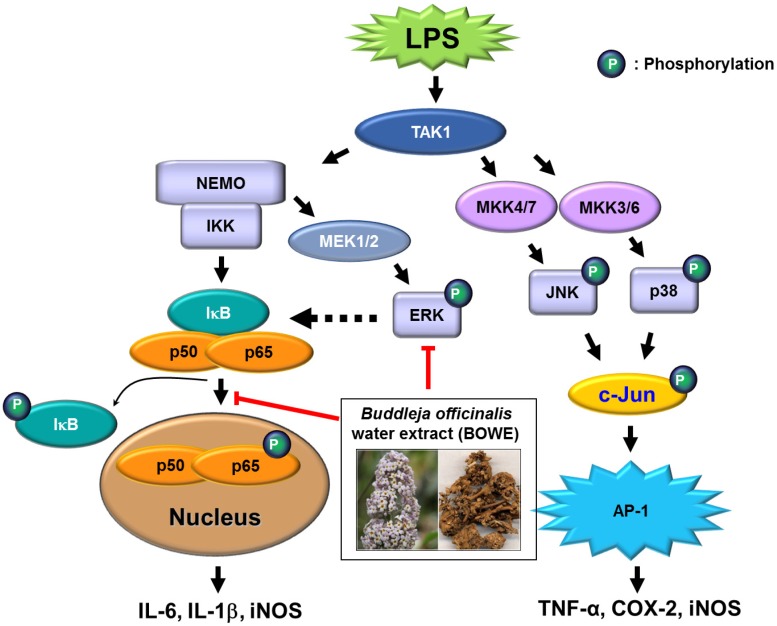
Buddleja officinalis has been traditionally used in the supportive treatment of inflammatory and neuronal diseases in Korea and China. Although several reports have shown the anti-inflammatory effects of Buddleja officinalis, the anti-neuroinflammatory effect has remained unclear. In this study, we aimed to investigate the inhibitory effects of flower buds of B. officinalis Maximowicz water extract (BOWE) on LPS-induced inflammatory processes in BV-2 microglial cells. BOWE dose-dependently inhibited the production of nitric oxide as well as iNOS mRNA expression. Moreover, BOWE prevented IL-1β and IL-6 mRNA expression. However, BOWE had no effect on LPS-induced COX-2 or TNF-a mRNA expression. The extract also had no effect on LPS-stimulated p38 MAPK, JNK, and c-Jun phosphorylation, whereas ERK1/2 phosphorylation was strongly inhibited by BOWE. BOWE also inhibited the LPS-induced degradation of IkB-α, and LPS-induced phosphorylation of p65 NF-kB protein. These data indicate that BOWE inhibited the nitric oxide production and pro-inflammatory gene expression in BV-2 microglial cells, possibly through a negative regulation of the NF-kB and ERK1/2 pathways. Further identification of the direct target molecule(s) of BOWE is required to support its use as an anti-neuroinflammatory agent against the neurodegenerative disorders.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
