

Top3β is an RNA topoisomerase that works with fragile X syndrome protein to promote synapse formation
- PMID: 23912945
- PMCID: PMC3853347
- DOI: 10.1038/nn.3479
Top3β is an RNA topoisomerase that works with fragile X syndrome protein to promote synapse formation
Abstract
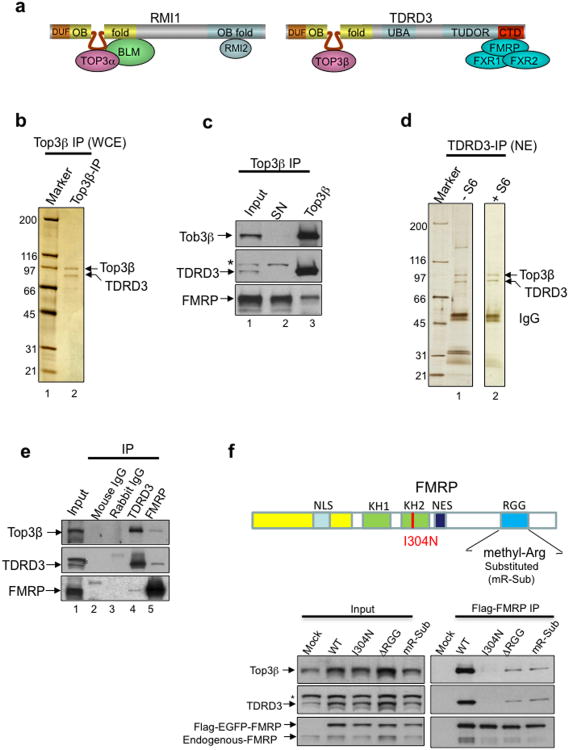
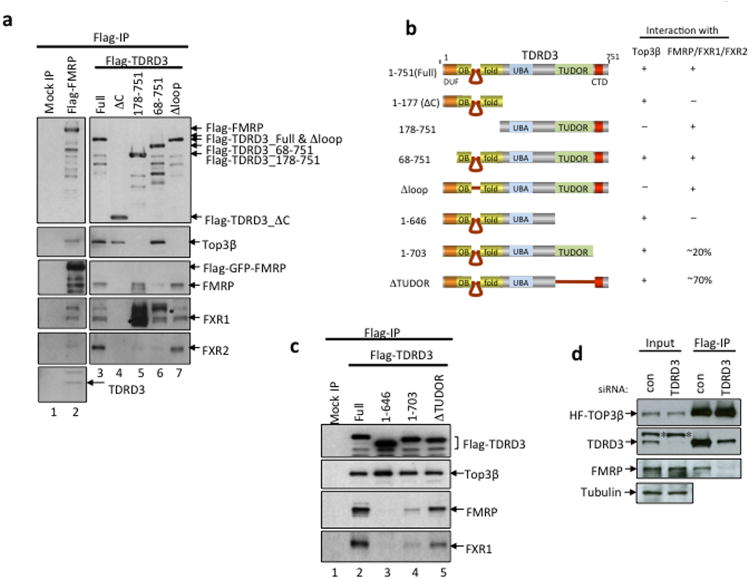
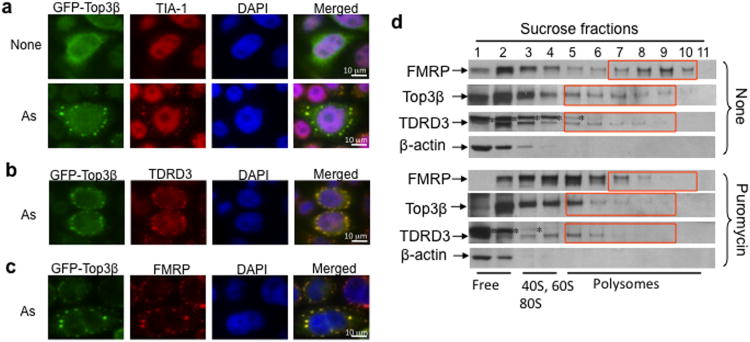
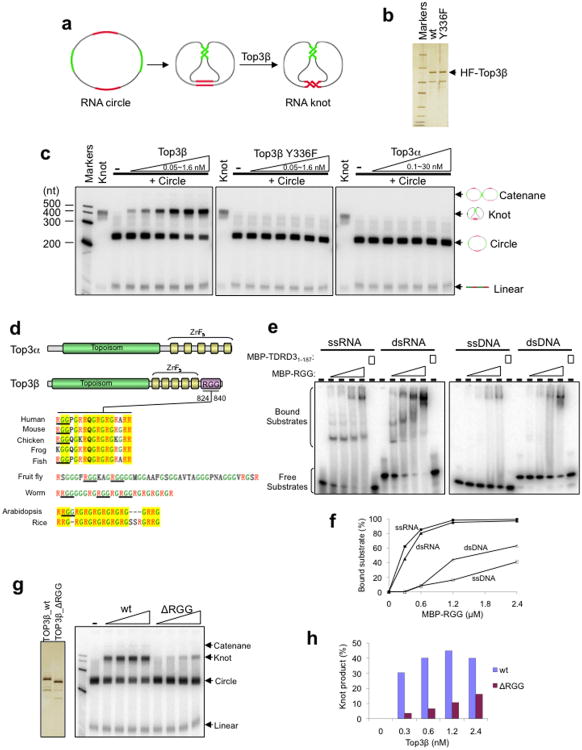
Topoisomerases are crucial for solving DNA topological problems, but they have not been linked to RNA metabolism. Here we show that human topoisomerase 3β (Top3β) is an RNA topoisomerase that biochemically and genetically interacts with FMRP, a protein that is deficient in fragile X syndrome and is known to regulate the translation of mRNAs that are important for neuronal function, abnormalities of which are linked to autism. Notably, the FMRP-Top3β interaction is abolished by a disease-associated mutation of FMRP, suggesting that Top3β may contribute to the pathogenesis of mental disorders. Top3β binds multiple mRNAs encoded by genes with neuronal functions linked to schizophrenia and autism. Expression of one such gene, that encoding protein tyrosine kinase 2 (ptk2, also known as focal adhesion kinase or FAK), is reduced in the neuromuscular junctions of Top3β mutant flies. Synapse formation is defective in Top3β mutant flies and mice, as well as in FMRP mutant flies and mice. Our findings suggest that Top3β acts as an RNA topoisomerase and works with FMRP to promote the expression of mRNAs that are crucial for neurodevelopment and mental health.
Figures

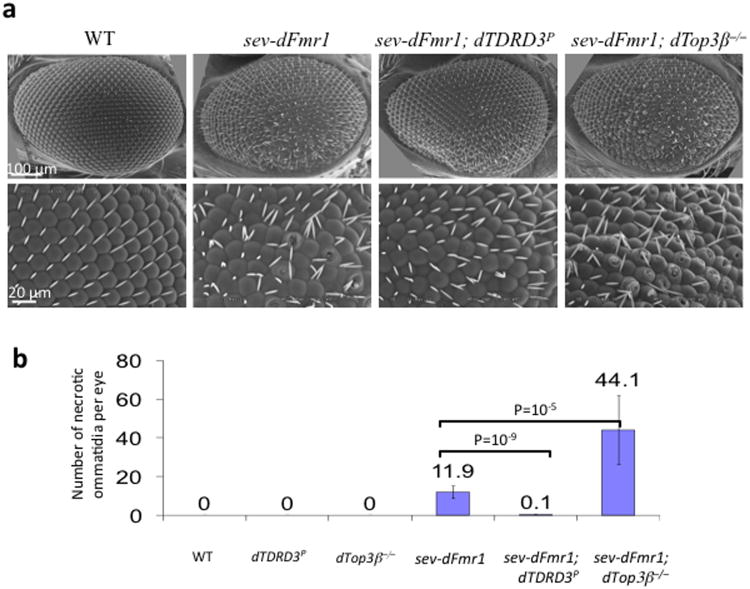
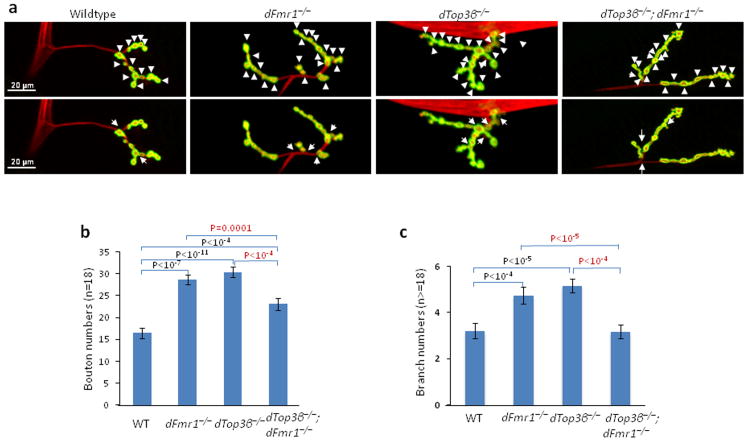
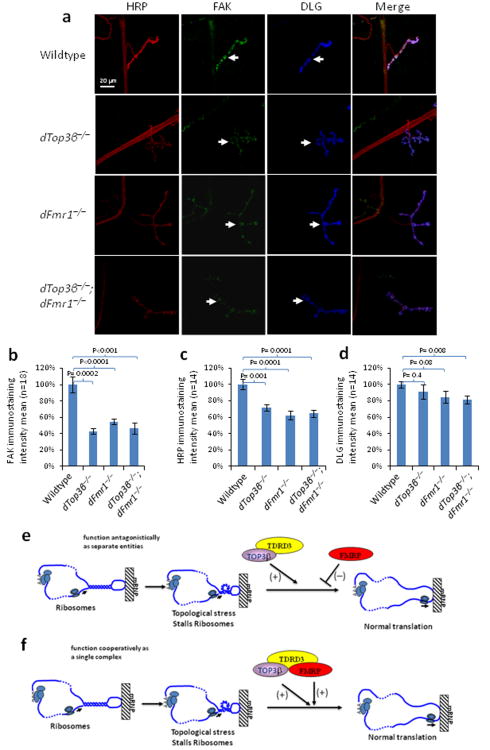
Comment in
-
The Top3β way to untangle RNA.Nat Neurosci. 2013 Sep;16(9):1163-4. doi: 10.1038/nn.3506. Nat Neurosci. 2013. PMID: 23982446 No abstract available.
-
Genetics: RNA topoisomerase involved in neurodevelopmental disorders.Nat Rev Neurol. 2013 Oct;9(10):542. doi: 10.1038/nrneurol.2013.178. Epub 2013 Sep 10. Nat Rev Neurol. 2013. PMID: 24018476 No abstract available.
References
-
- Wang JC. Cellular roles of DNA topoisomerases: a molecular perspective. Nat Rev Mol Cell Biol. 2002;3:430–440. - PubMed
-
- Plank JL, Chu SH, Pohlhaus JR, Wilson-Sali T, Hsieh TS. Drosophila melanogaster topoisomerase IIIalpha preferentially relaxes a positively or negatively supercoiled bubble substrate and is essential during development. J Biol Chem. 2005;280:3564–3573. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
