

Addiction is a Reward Deficit and Stress Surfeit Disorder
- PMID: 23914176
- PMCID: PMC3730086
- DOI: 10.3389/fpsyt.2013.00072
Addiction is a Reward Deficit and Stress Surfeit Disorder
Abstract
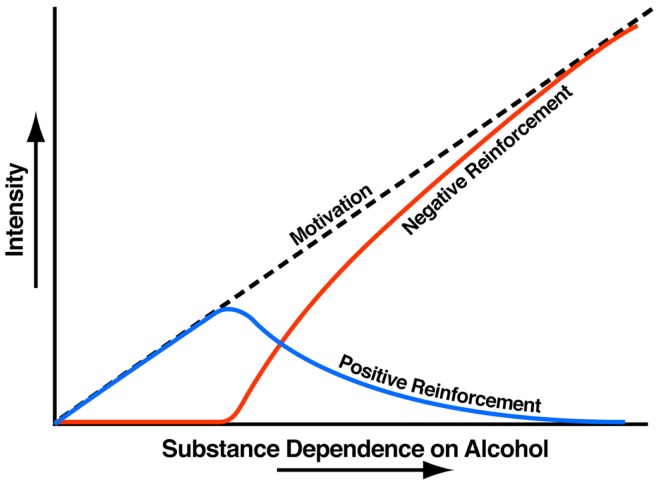
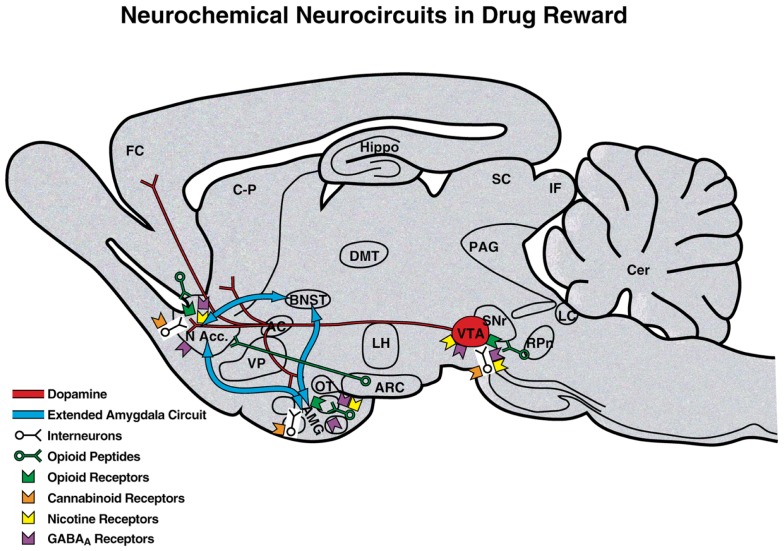
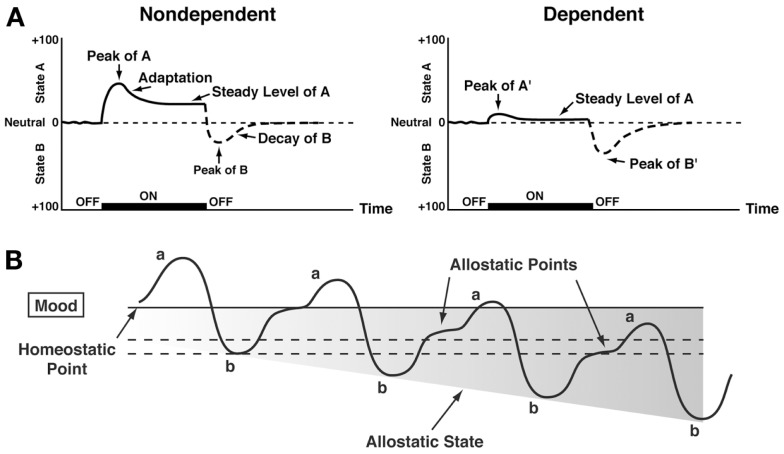
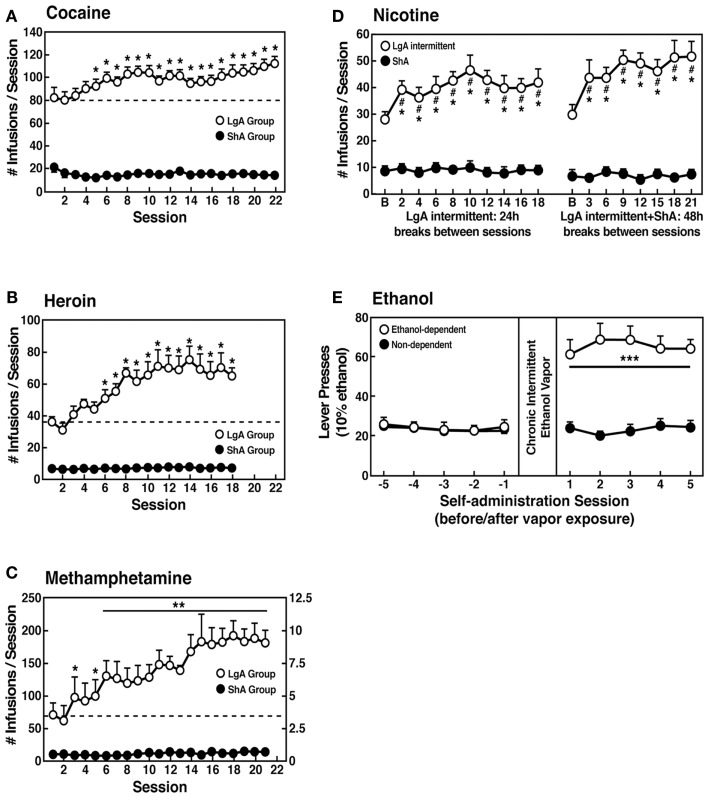
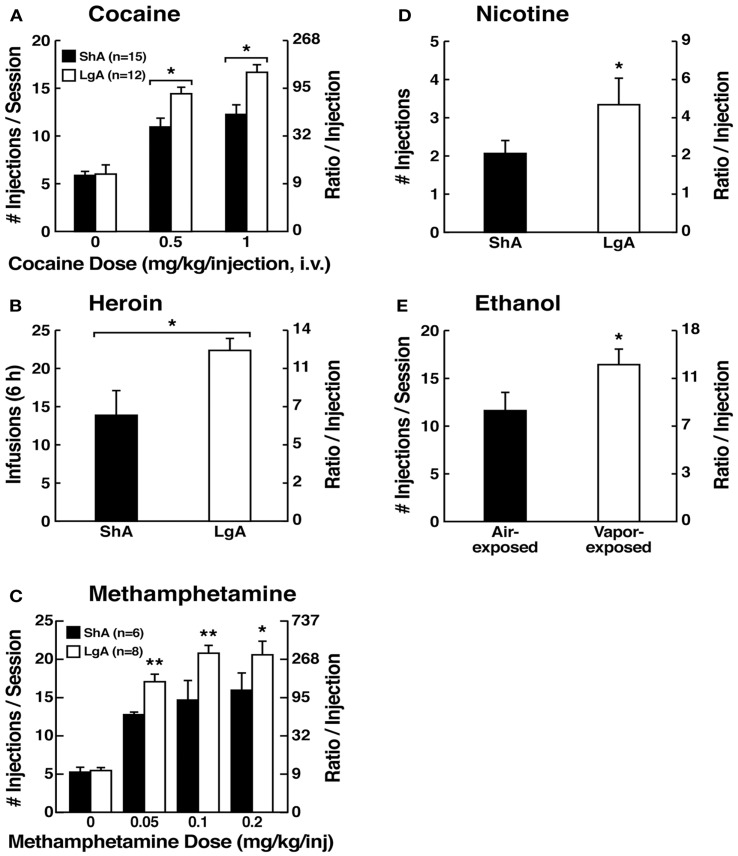
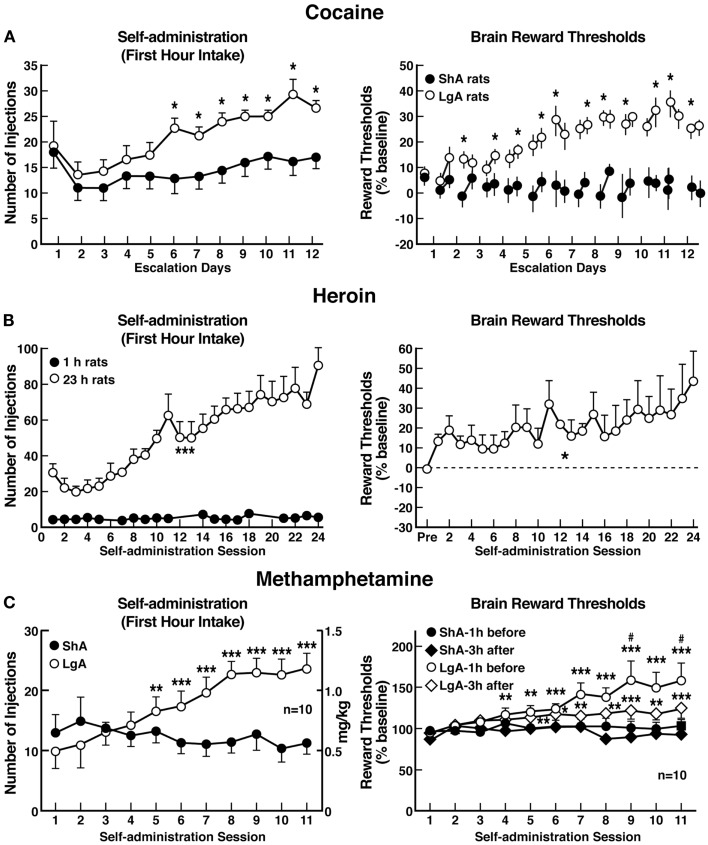
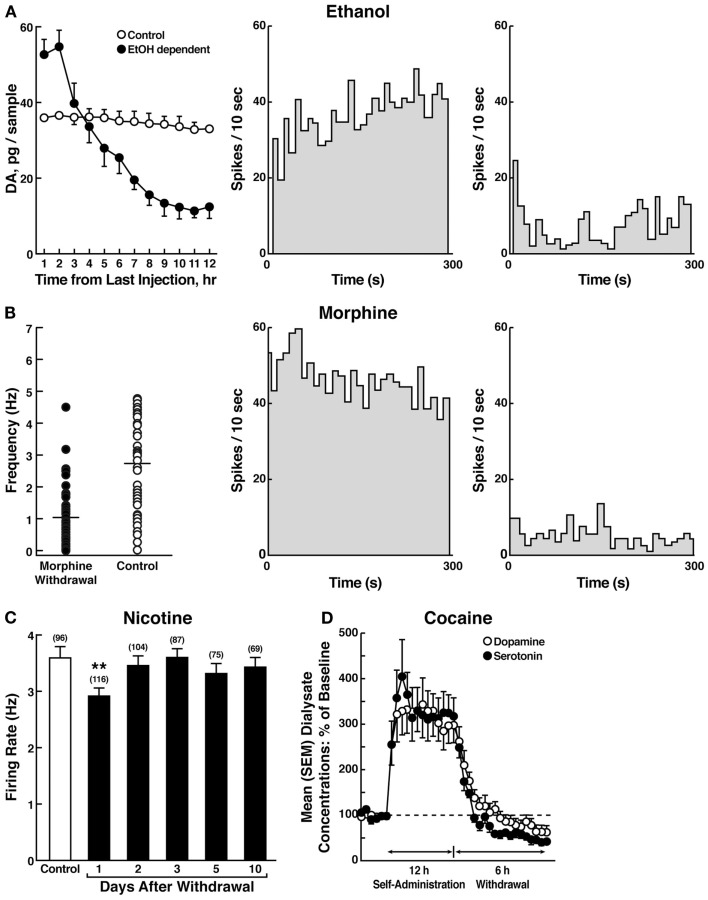
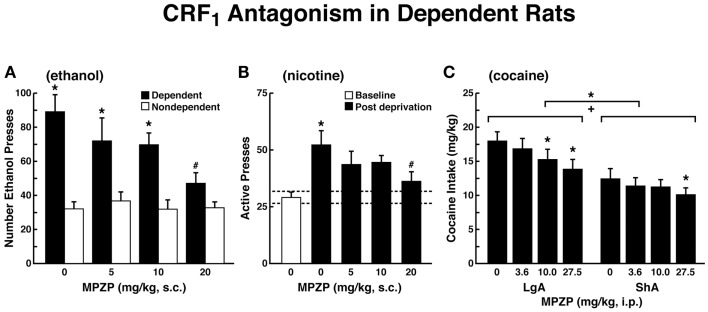
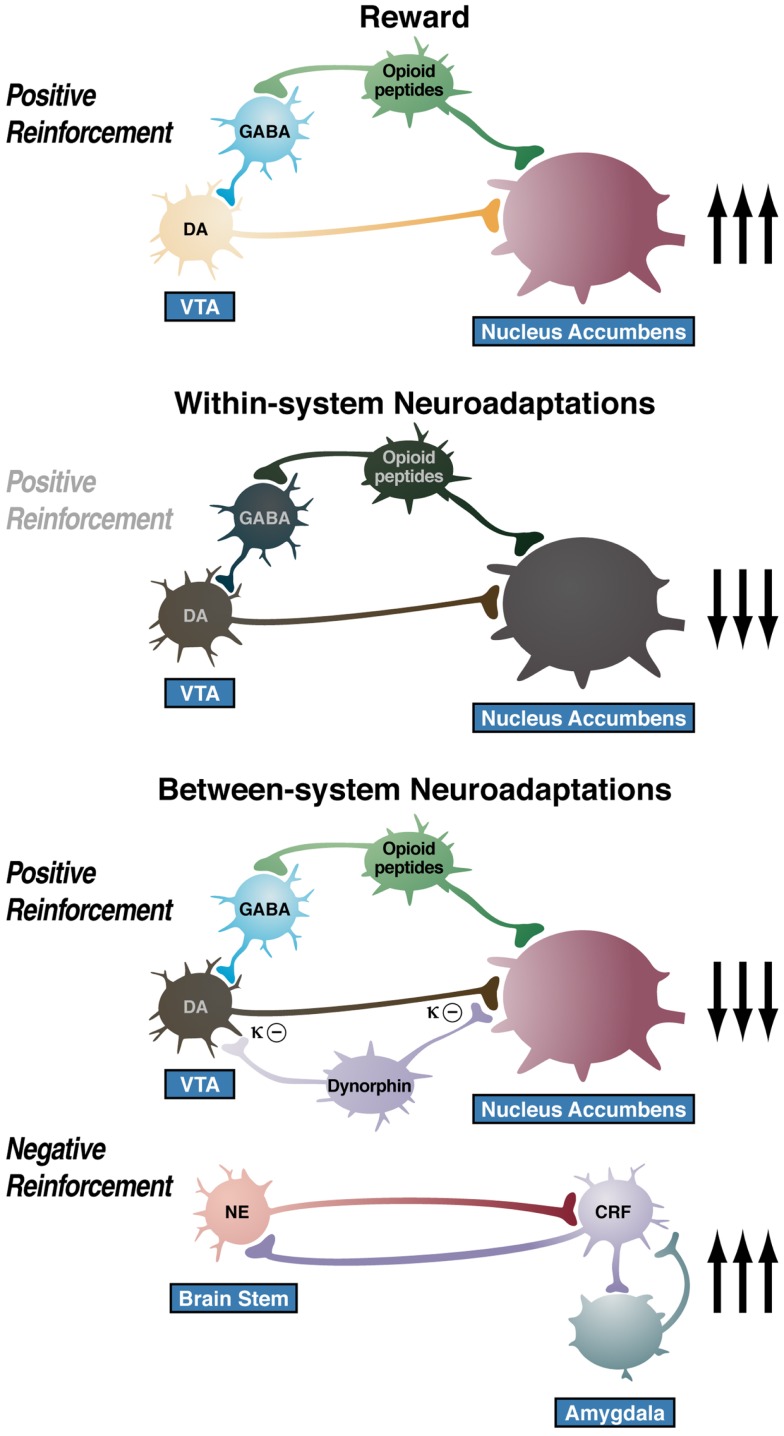
Drug addiction can be defined by a three-stage cycle - binge/intoxication, withdrawal/negative affect, and preoccupation/anticipation - that involves allostatic changes in the brain reward and stress systems. Two primary sources of reinforcement, positive and negative reinforcement, have been hypothesized to play a role in this allostatic process. The negative emotional state that drives negative reinforcement is hypothesized to derive from dysregulation of key neurochemical elements involved in the brain reward and stress systems. Specific neurochemical elements in these structures include not only decreases in reward system function (within-system opponent processes) but also recruitment of the brain stress systems mediated by corticotropin-releasing factor (CRF) and dynorphin-κ opioid systems in the ventral striatum, extended amygdala, and frontal cortex (both between-system opponent processes). CRF antagonists block anxiety-like responses associated with withdrawal, block increases in reward thresholds produced by withdrawal from drugs of abuse, and block compulsive-like drug taking during extended access. Excessive drug taking also engages the activation of CRF in the medial prefrontal cortex, paralleled by deficits in executive function that may facilitate the transition to compulsive-like responding. Neuropeptide Y, a powerful anti-stress neurotransmitter, has a profile of action on compulsive-like responding for ethanol similar to a CRF1 antagonist. Blockade of the κ opioid system can also block dysphoric-like effects associated with withdrawal from drugs of abuse and block the development of compulsive-like responding during extended access to drugs of abuse, suggesting another powerful brain stress system that contributes to compulsive drug seeking. The loss of reward function and recruitment of brain systems provide a powerful neurochemical basis that drives the compulsivity of addiction.
Keywords: compulsive; corticotropin-releasing factor; dynorphin; extended amygdala; opponent process; prefrontal cortex; reward; withdrawal.
Figures
References
-
- Simon H, Stinus L, Tassin JP, Lavielle S, Blanc G, Thierry AM, et al. Is the dopaminergic mesocorticolimbic system necessary for intracranial self-stimulation? Biochemical and behavioral studies from A10 cell bodies and terminals. Behav Neural Biol (1979) 27:125–4510.1016/S0163-1047(79)91745-X - DOI - PubMed
-
- Kornetsky C, Esposito RU. Euphorigenic drugs: effects on the reward pathways of the brain. Fed Proc (1979) 38:2473–6 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical