

The hippocampus, time, and memory across scales
- PMID: 23915126
- PMCID: PMC3982793
- DOI: 10.1037/a0033621
The hippocampus, time, and memory across scales
Abstract
A wealth of experimental studies with animals have offered insights about how neural networks within the hippocampus support the temporal organization of memories. These studies have revealed the existence of "time cells" that encode moments in time, much as the well-known "place cells" map locations in space. Another line of work inspired by human behavioral studies suggests that episodic memories are mediated by a state of temporal context that changes gradually over long time scales, up to at least a few thousand seconds. In this view, the "mental time travel" hypothesized to support the experience of episodic memory corresponds to a "jump back in time" in which a previous state of temporal context is recovered. We suggest that these 2 sets of findings could be different facets of a representation of temporal history that maintains a record at the last few thousand seconds of experience. The ability to represent long time scales comes at the cost of discarding precise information about when a stimulus was experienced--this uncertainty becomes greater for events further in the past. We review recent computational work that describes a mechanism that could construct such a scale-invariant representation. Taken as a whole, this suggests the hippocampus plays its role in multiple aspects of cognition by representing events embedded in a general spatiotemporal context. The representation of internal time can be useful across nonhippocampal memory systems.
PsycINFO Database Record (c) 2013 APA, all rights reserved.
Figures
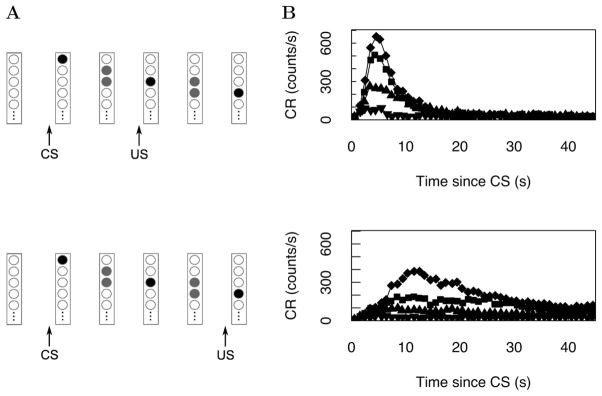
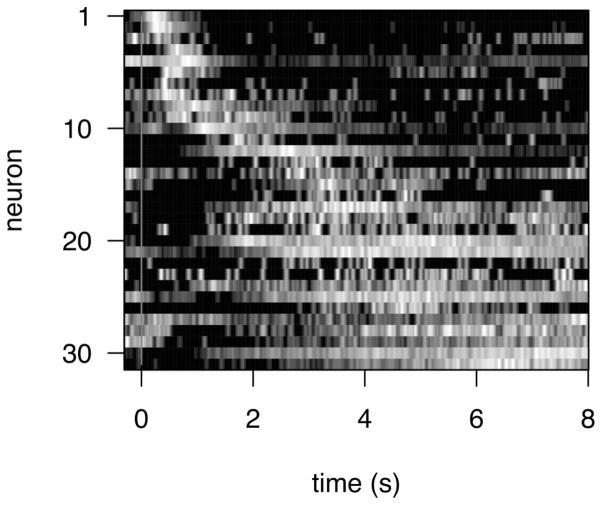
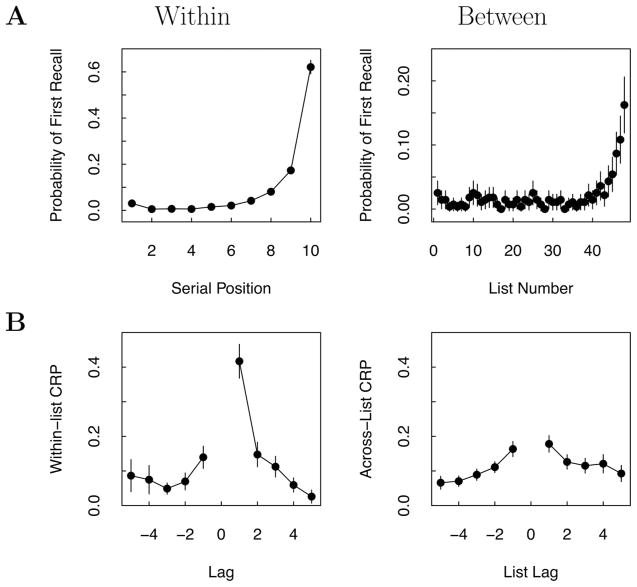
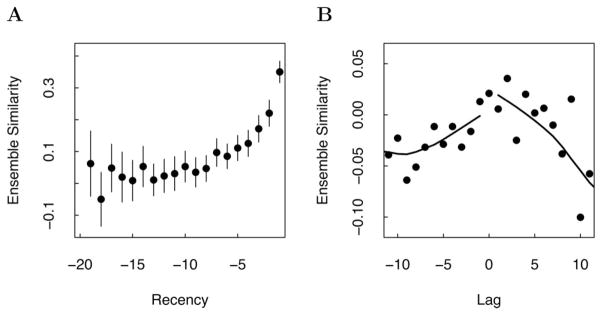

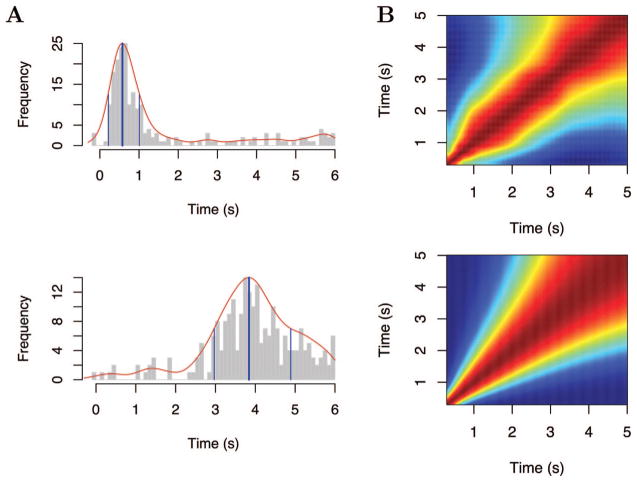
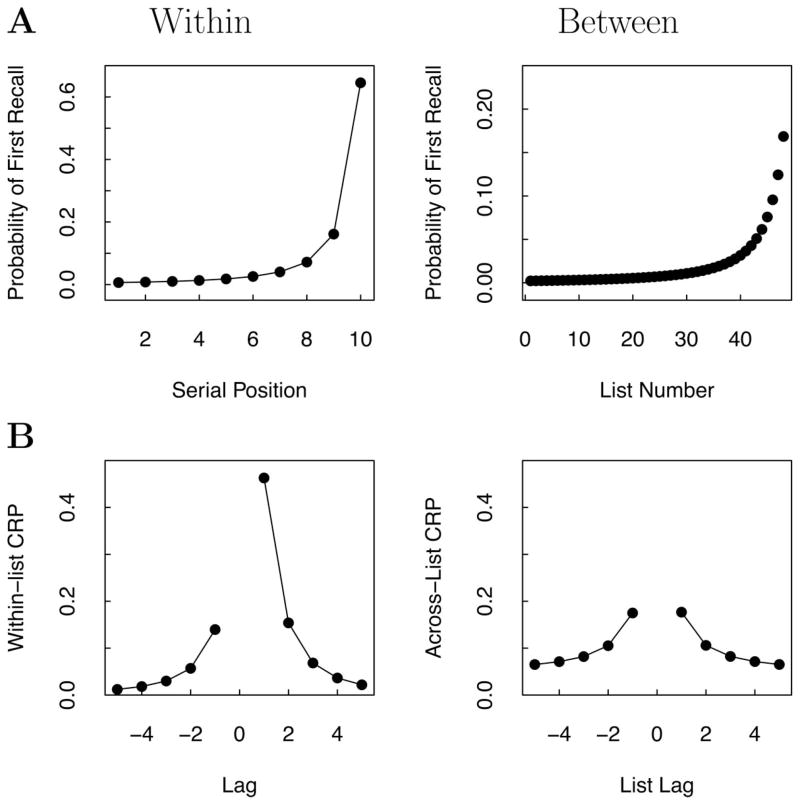
References
-
- Anderson J, Schooler L. Reflections of the environment in memory. Psychological Science. 1991;2:396–408. doi: 10.1111/j.1467-9280.1991.tb00174.x. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
