

Bacterial cheating drives the population dynamics of cooperative antibiotic resistance plasmids
- PMID: 23917989
- PMCID: PMC3779801
- DOI: 10.1038/msb.2013.39
Bacterial cheating drives the population dynamics of cooperative antibiotic resistance plasmids
Abstract
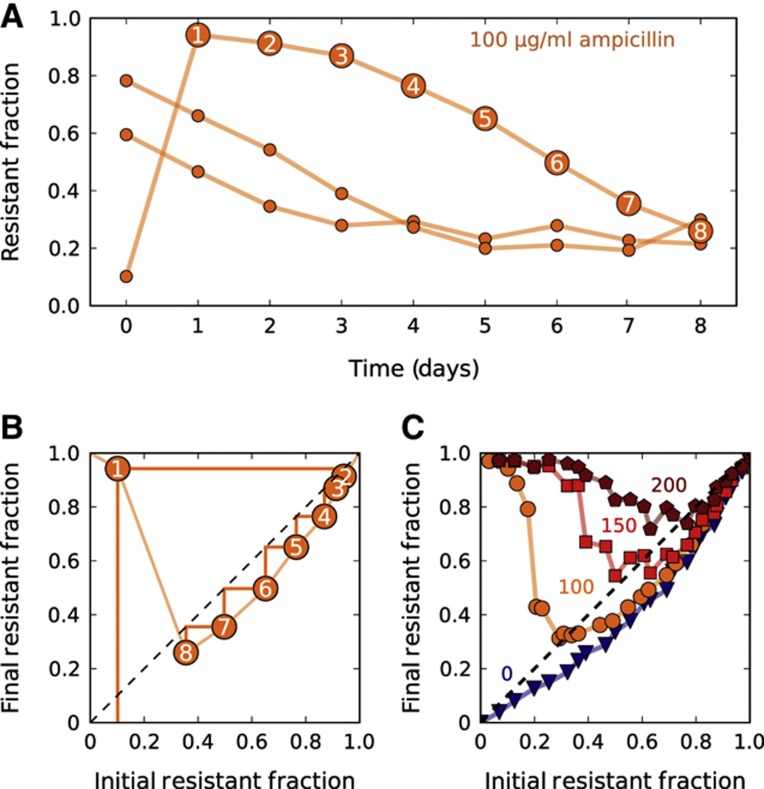
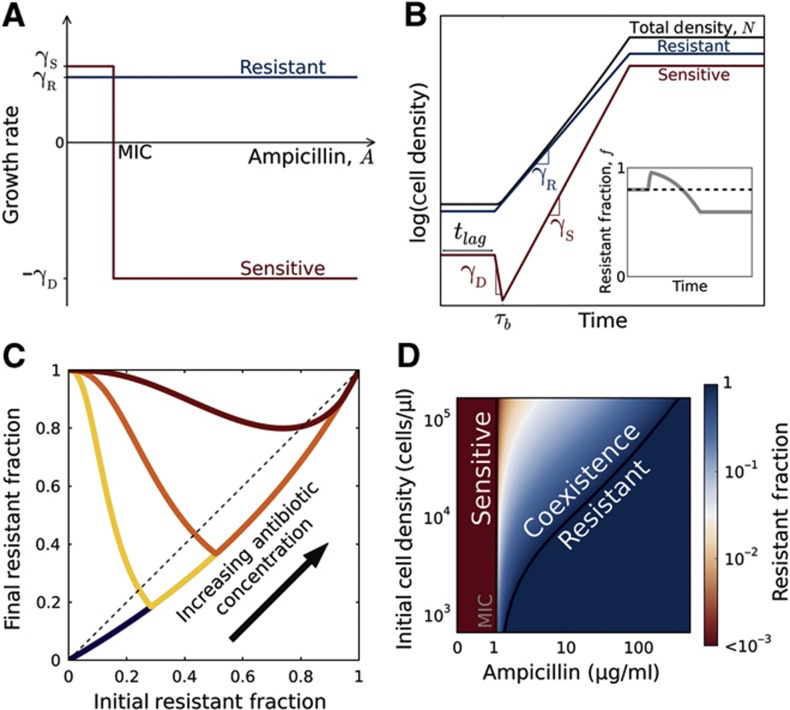
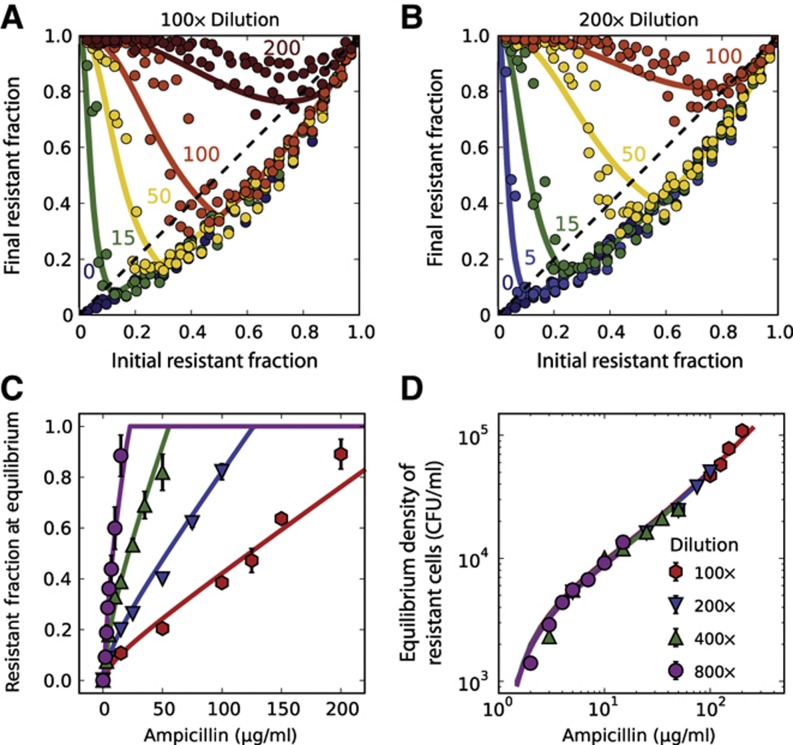
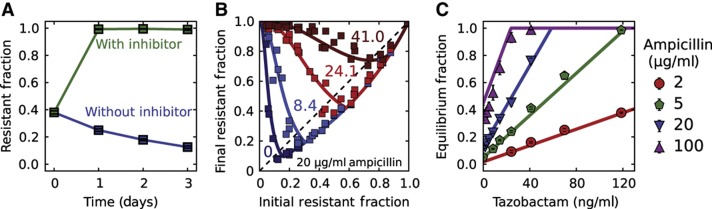
Inactivation of β-lactam antibiotics by resistant bacteria is a 'cooperative' behavior that may allow sensitive bacteria to survive antibiotic treatment. However, the factors that determine the fraction of resistant cells in the bacterial population remain unclear, indicating a fundamental gap in our understanding of how antibiotic resistance evolves. Here, we experimentally track the spread of a plasmid that encodes a β-lactamase enzyme through the bacterial population. We find that independent of the initial fraction of resistant cells, the population settles to an equilibrium fraction proportional to the antibiotic concentration divided by the cell density. A simple model explains this behavior, successfully predicting a data collapse over two orders of magnitude in antibiotic concentration. This model also successfully predicts that adding a commonly used β-lactamase inhibitor will lead to the spread of resistance, highlighting the need to incorporate social dynamics into the study of antibiotic resistance.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Andersson DI (2006) The biological cost of mutational antibiotic resistance: any practical conclusions? Curr Opin Microbiol 9: 461–465 - PubMed
-
- Bonomo RA, Tolmasky ME (eds) (2007) Enzyme-Mediated Resistance to Antibiotics: Mechanisms, Dissemination, and Prospects for Inhibition 1st edn. Amer Society for Microbiology: Washington, DC,
-
- Bouma JE, Lenski RE (1988) Evolution of a bacteria/plasmid association. Nature 335: 351–352 - PubMed
-
- Brook I (1984) The role of β-lactamase-producing bacteria in the persistence of streptococcal tonsillar infection. Clin Infect Dis 6: 601–607 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
