

Conditionals by inversion provide a universal method for the generation of conditional alleles
- PMID: 23918385
- PMCID: PMC3752204
- DOI: 10.1073/pnas.1217812110
Conditionals by inversion provide a universal method for the generation of conditional alleles
Abstract
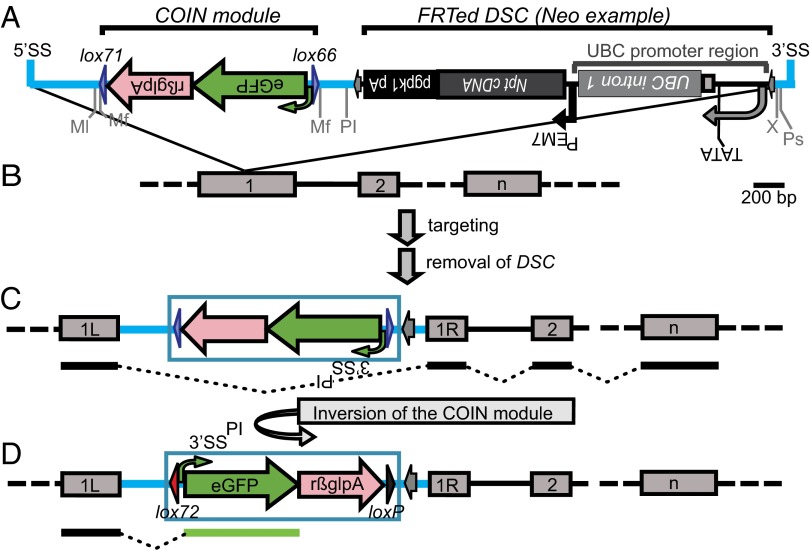
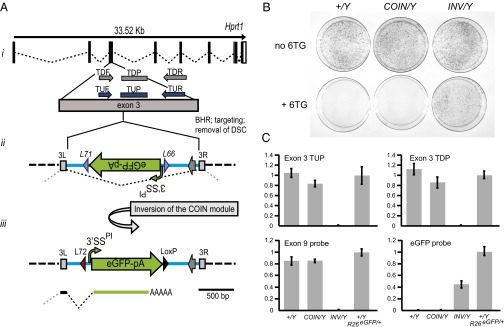
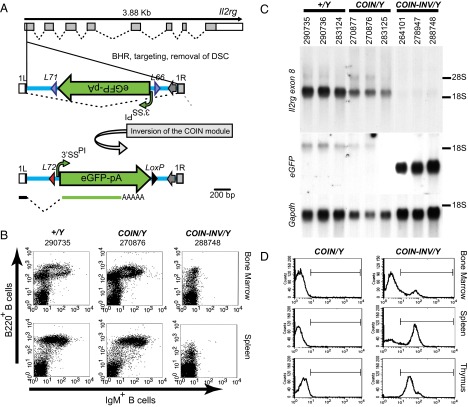
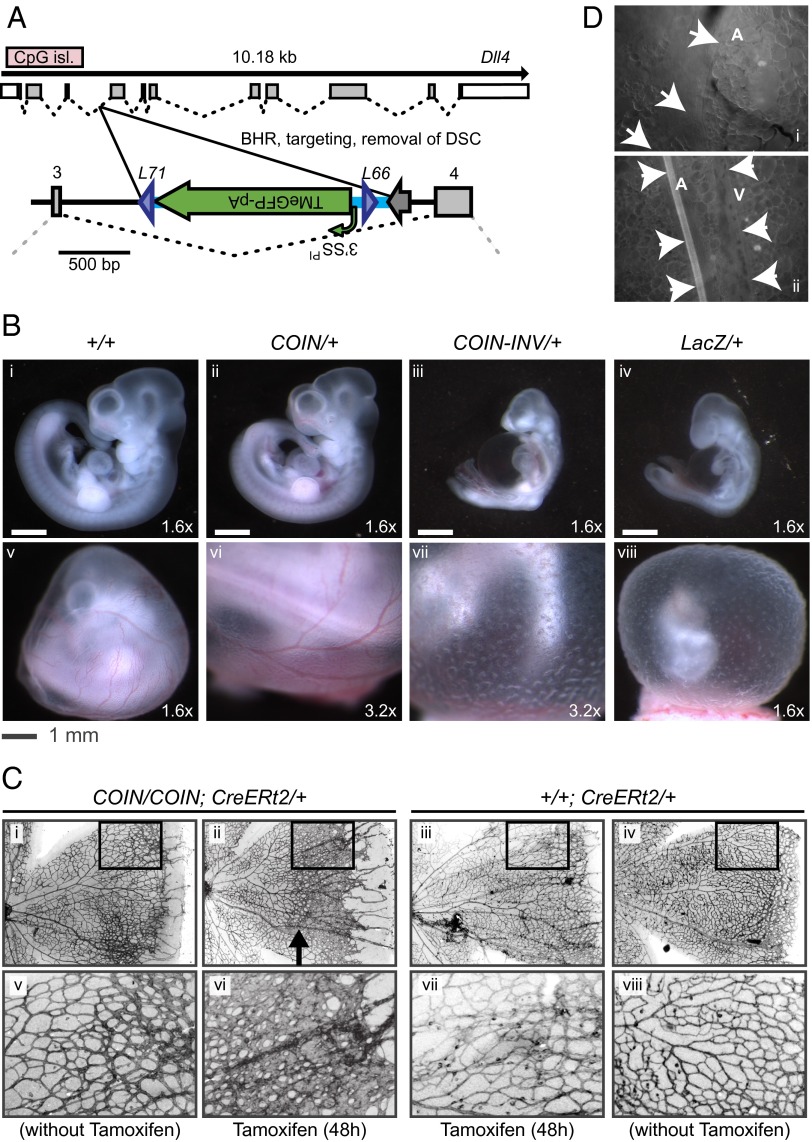
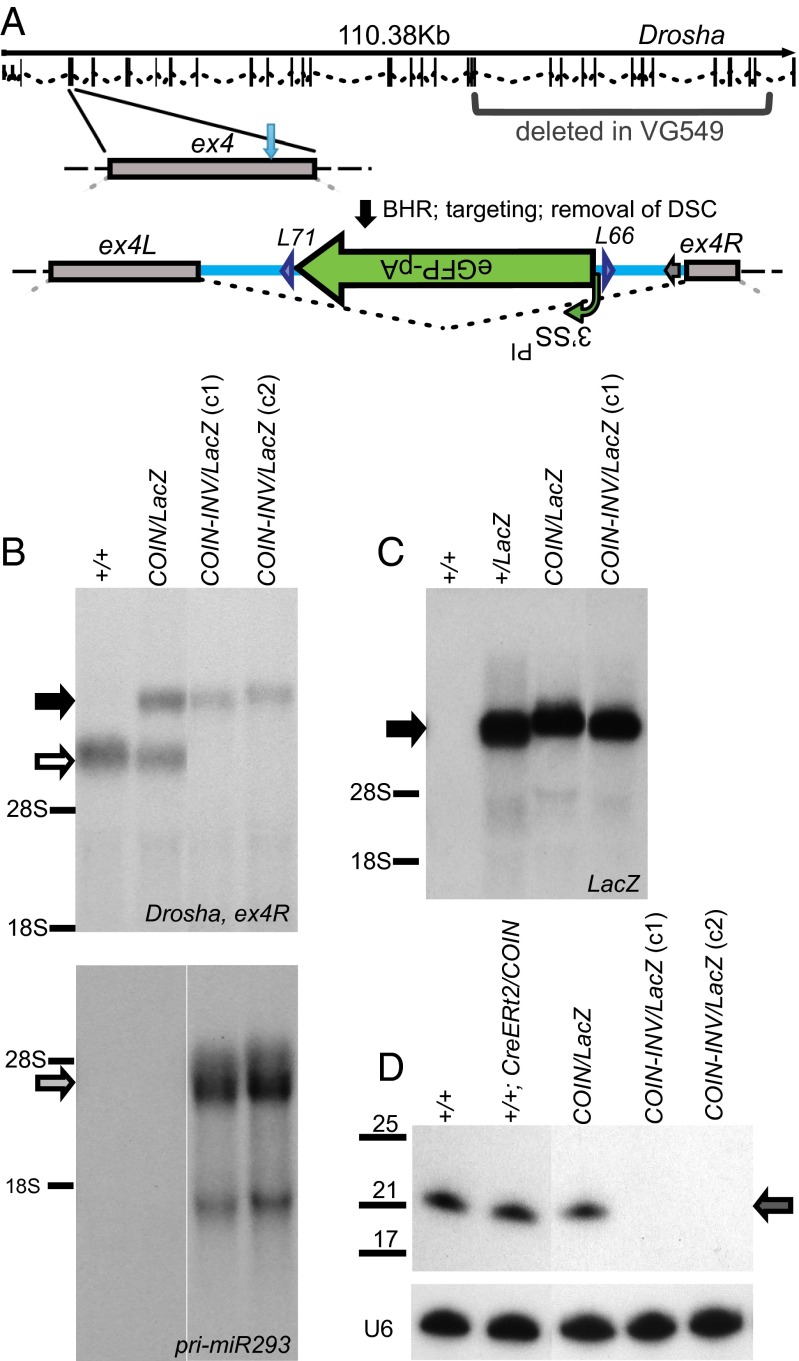
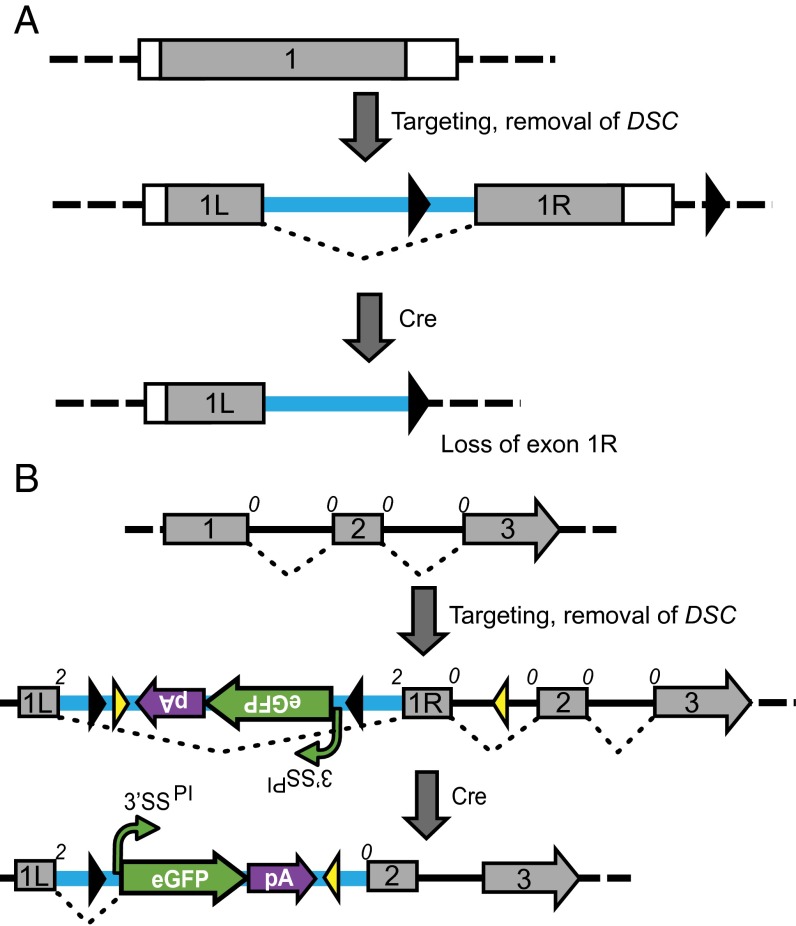
Conditional mutagenesis is becoming a method of choice for studying gene function, but constructing conditional alleles is often laborious, limited by target gene structure, and at times, prone to incomplete conditional ablation. To address these issues, we developed a technology termed conditionals by inversion (COIN). Before activation, COINs contain an inverted module (COIN module) that lies inertly within the antisense strand of a resident gene. When inverted into the sense strand by a site-specific recombinase, the COIN module causes termination of the target gene's transcription and simultaneously provides a reporter for tracking this event. COIN modules can be inserted into natural introns (intronic COINs) or directly into coding exons as part of an artificial intron (exonic COINs), greatly simplifying allele design and increasing flexibility over previous conditional KO approaches. Detailed analysis of over 20 COIN alleles establishes the reliability of the method and its broad applicability to any gene, regardless of exon-intron structure. Our extensive testing provides rules that help ensure success of this approach and also explains why other currently available conditional approaches often fail to function optimally. Finally, the ability to split exons using the COIN's artificial intron opens up engineering modalities for the generation of multifunctional alleles.
Keywords: conditional-null; genome engineering.
Conflict of interest statement
All authors of this manuscript are employed by Regeneron Pharmaceuticals, Inc., and some of the authors own a substantial amount of Regeneron stock.
Figures
References
-
- Branda CS, Dymecki SM. Talking about a revolution: The impact of site-specific recombinases on genetic analyses in mice. Dev Cell. 2004;6(1):7–28. - PubMed
-
- Testa G, et al. A reliable lacZ expression reporter cassette for multipurpose, knockout-first alleles. Genesis. 2004;38(3):151–158. - PubMed
-
- Schnütgen F, et al. A directional strategy for monitoring Cre-mediated recombination at the cellular level in the mouse. Nat Biotechnol. 2003;21(5):562–565. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
