

Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system
- PMID: 23918387
- PMCID: PMC3752207
- DOI: 10.1073/pnas.1308335110
Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system
Abstract
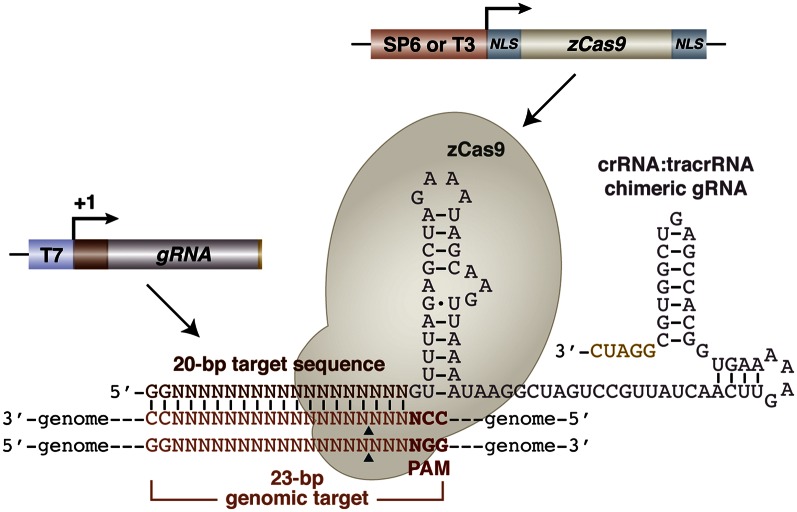
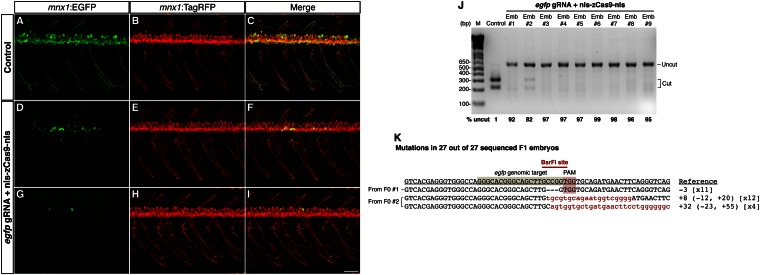
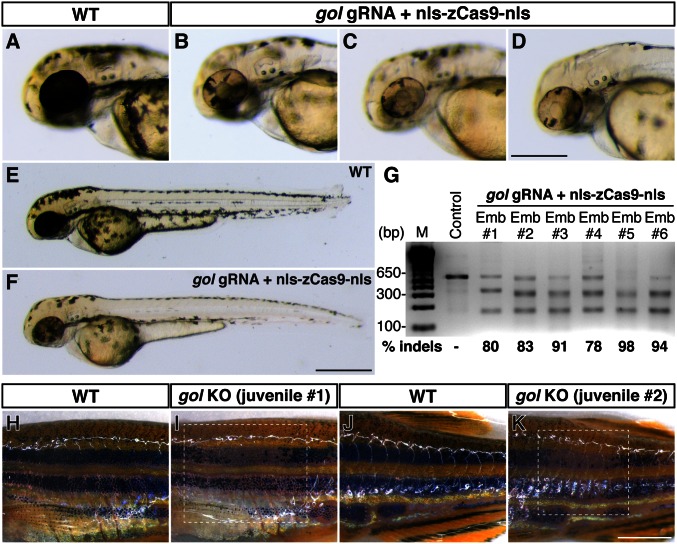
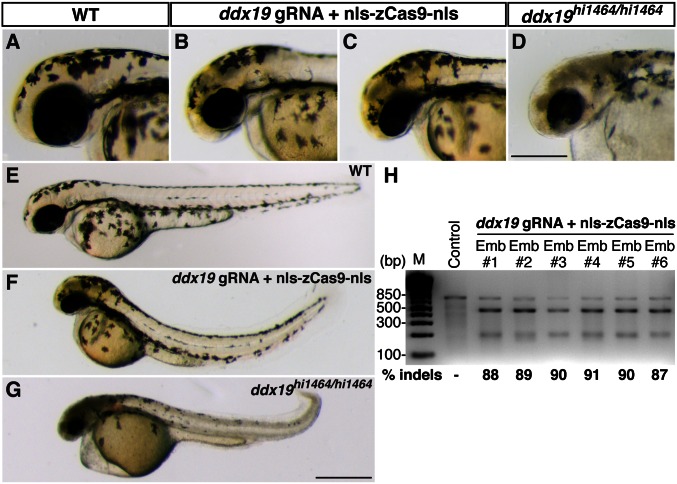
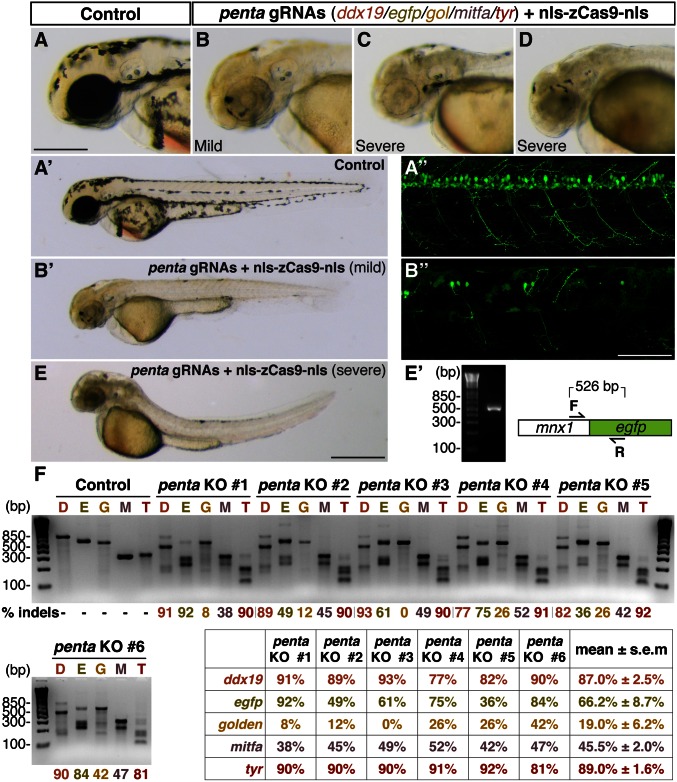
A simple and robust method for targeted mutagenesis in zebrafish has long been sought. Previous methods generate monoallelic mutations in the germ line of F0 animals, usually delaying homozygosity for the mutation to the F2 generation. Generation of robust biallelic mutations in the F0 would allow for phenotypic analysis directly in injected animals. Recently the type II prokaryotic clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated proteins (Cas) system has been adapted to serve as a targeted genome mutagenesis tool. Here we report an improved CRISPR/Cas system in zebrafish with custom guide RNAs and a zebrafish codon-optimized Cas9 protein that efficiently targeted a reporter transgene Tg(-5.1mnx1:egfp) and four endogenous loci (tyr, golden, mitfa, and ddx19). Mutagenesis rates reached 75-99%, indicating that most cells contained biallelic mutations. Recessive null-like phenotypes were observed in four of the five targeting cases, supporting high rates of biallelic gene disruption. We also observed efficient germ-line transmission of the Cas9-induced mutations. Finally, five genomic loci can be targeted simultaneously, resulting in multiple loss-of-function phenotypes in the same injected fish. This CRISPR/Cas9 system represents a highly effective and scalable gene knockout method in zebrafish and has the potential for applications in other model organisms.
Keywords: RNA-guided mutagenesis; genome engineering; pigmentation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Urnov FD, et al. Highly efficient endogenous human gene correction using designed zinc-finger nucleases. Nature. 2005;435(7042):646–651. - PubMed
-
- Bhaya D, Davison M, Barrangou R. CRISPR-Cas systems in bacteria and archaea: Versatile small RNAs for adaptive defense and regulation. Annu Rev Genet. 2011;45:273–297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
