

AMP-activated protein kinase regulates nicotinamide phosphoribosyl transferase expression in skeletal muscle
- PMID: 23918774
- PMCID: PMC3810819
- DOI: 10.1113/jphysiol.2013.259515
AMP-activated protein kinase regulates nicotinamide phosphoribosyl transferase expression in skeletal muscle
Abstract
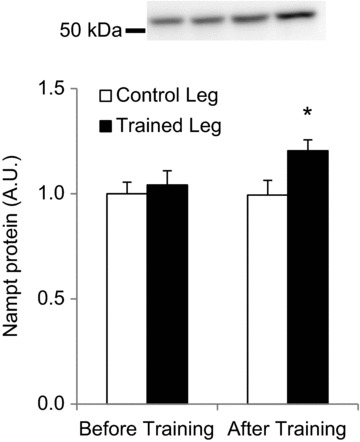
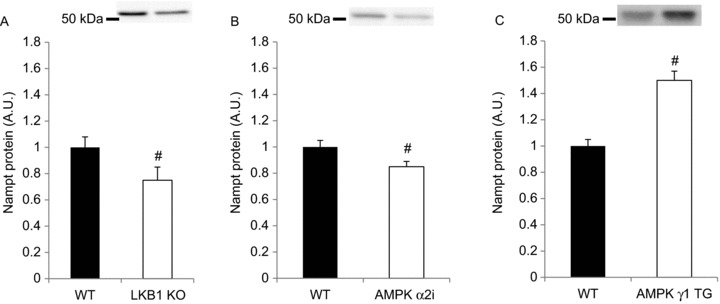
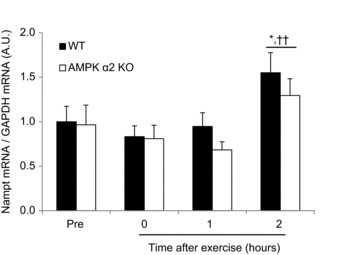
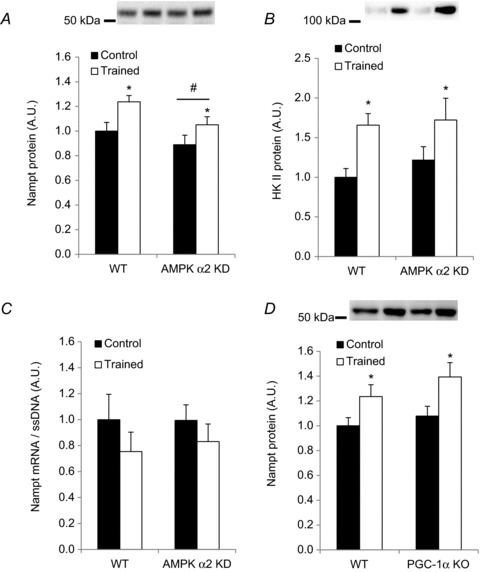
Deacetylases such as sirtuins (SIRTs) convert NAD to nicotinamide (NAM). Nicotinamide phosphoribosyl transferase (Nampt) is the rate-limiting enzyme in the NAD salvage pathway responsible for converting NAM to NAD to maintain cellular redox state. Activation of AMP-activated protein kinase (AMPK) increases SIRT activity by elevating NAD levels. As NAM directly inhibits SIRTs, increased Nampt activation or expression could be a metabolic stress response. Evidence suggests that AMPK regulates Nampt mRNA content, but whether repeated AMPK activation is necessary for increasing Nampt protein levels is unknown. To this end, we assessed whether exercise training- or 5-amino-1-β-D-ribofuranosyl-imidazole-4-carboxamide (AICAR)-mediated increases in skeletal muscle Nampt abundance are AMPK dependent. One-legged knee-extensor exercise training in humans increased Nampt protein by 16% (P < 0.05) in the trained, but not the untrained leg. Moreover, increases in Nampt mRNA following acute exercise or AICAR treatment (P < 0.05 for both) were maintained in mouse skeletal muscle lacking a functional AMPK α2 subunit. Nampt protein was reduced in skeletal muscle of sedentary AMPK α2 kinase dead (KD), but 6.5 weeks of endurance exercise training increased skeletal muscle Nampt protein to a similar extent in both wild-type (WT) (24%) and AMPK α2 KD (18%) mice. In contrast, 4 weeks of daily AICAR treatment increased Nampt protein in skeletal muscle in WT mice (27%), but this effect did not occur in AMPK α2 KD mice. In conclusion, functional α2-containing AMPK heterotrimers are required for elevation of skeletal muscle Nampt protein, but not mRNA induction. These findings suggest AMPK plays a post-translational role in the regulation of skeletal muscle Nampt protein abundance, and further indicate that the regulation of cellular energy charge and nutrient sensing is mechanistically related.
Figures
References
-
- Andersen P, Adams RP, Sjøgaard G, Thorboe A, Saltin B. Dynamic knee extension as model for study of isolated exercising muscle in humans. J Appl Physiol. 1985;59:1647–1653. - PubMed
-
- Bailey CJ, Puah JA. Effect of metformin on glucose metabolism in mouse soleus muscle. Diabete Metab. 1986;12:212–218. - PubMed
-
- Barnes BR, Glund S, Long YC, Hjälm G, Andersson L, Zierath JR. 5′-AMP-activated protein kinase regulates skeletal muscle glycogen content and ergogenics. FASEB J. 2005;19:773–779. - PubMed
-
- Barnes BR, Marklund S, Steiler TL, Walter M, Hjälm G, Amarger V, Mahlapuu M, Leng Y, Johansson C, Galuska D, Lindgren K, Abrink M, Stapleton D, Zierath JR, Andersson L. The 5′-AMP-activated protein kinase gamma3 isoform has a key role in carbohydrate and lipid metabolism in glycolytic skeletal muscle. J Biol Chem. 2004;279:38441–38447. - PubMed
-
- Barré L, Richardson C, Hirshman MF, Brozinick J, Fiering S, Kemp BE, Goodyear LJ, Witters LA. Genetic model for the chronic activation of skeletal muscle AMP-activated protein kinase leads to glycogen accumulation. Am J Physiol Endocrinol Metab. 2007;292:E802–E811. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous