

Towards an integrated experimental-theoretical approach for assessing the mechanistic basis of hair and feather morphogenesis
- PMID: 23919127
- PMCID: PMC3363042
- DOI: 10.1098/rsfs.2011.0122
Towards an integrated experimental-theoretical approach for assessing the mechanistic basis of hair and feather morphogenesis
Abstract
In his seminal 1952 paper, 'The Chemical Basis of Morphogenesis', Alan Turing lays down a milestone in the application of theoretical approaches to understand complex biological processes. His deceptively simple demonstration that a system of reacting and diffusing chemicals could, under certain conditions, generate spatial patterning out of homogeneity provided an elegant solution to the problem of how one of nature's most intricate events occurs: the emergence of structure and form in the developing embryo. The molecular revolution that has taken place during the six decades following this landmark publication has now placed this generation of theoreticians and biologists in an excellent position to rigorously test the theory and, encouragingly, a number of systems have emerged that appear to conform to some of Turing's fundamental ideas. In this paper, we describe the history and more recent integration between experiment and theory in one of the key models for understanding pattern formation: the emergence of feathers and hair in the skins of birds and mammals.
Keywords: Turing patterns; activator–inhibitor; feather buds and hair follicles; morphogenesis; reaction–diffusion; skin patterning.
Figures

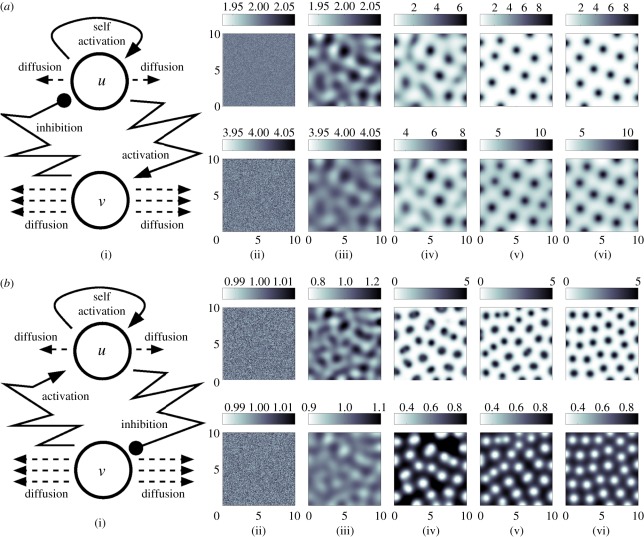
 and
and  with d = 0.025, a = 0.5. (b(i)) Schematic of the underlying interactions in a cross activator–inhibitor system. (b(ii)–b(vi)) Simulation of a cross AI system showing the spatial pattern for (top) u and (bottom) v at (b(ii)) t = 0, (b(iii)) t = 10, (b(iv)) t = 20, (b(v)) t = 50, at (b(vi)) t = 1000. Peaks of u and v concentration lie out of phase. Simulations were performed using Schnakenberg [43] kinetics,
with d = 0.025, a = 0.5. (b(i)) Schematic of the underlying interactions in a cross activator–inhibitor system. (b(ii)–b(vi)) Simulation of a cross AI system showing the spatial pattern for (top) u and (bottom) v at (b(ii)) t = 0, (b(iii)) t = 10, (b(iv)) t = 20, (b(v)) t = 50, at (b(vi)) t = 1000. Peaks of u and v concentration lie out of phase. Simulations were performed using Schnakenberg [43] kinetics,  and
and  , with d = 0.025, a = 1.0. For both sets of simulations, initial conditions were set at a small perturbation of the uniform steady state and zero-flux conditions were employed at the boundaries of a square field of dimensions 10 × 10.
, with d = 0.025, a = 1.0. For both sets of simulations, initial conditions were set at a small perturbation of the uniform steady state and zero-flux conditions were employed at the boundaries of a square field of dimensions 10 × 10.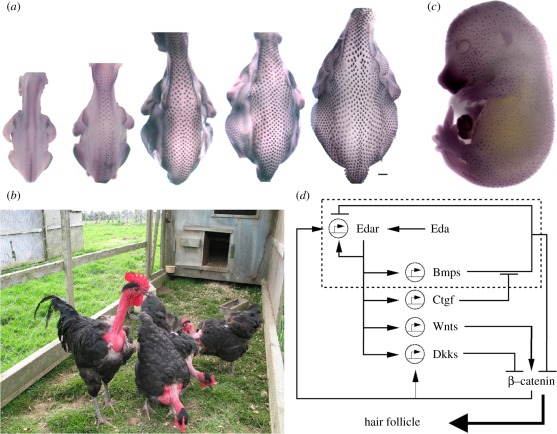
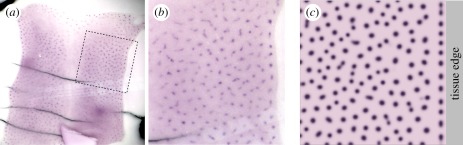
 and solved on a square field of dimension 3 × 3. For experimental details, see the study of Mou et al. [26].
and solved on a square field of dimension 3 × 3. For experimental details, see the study of Mou et al. [26].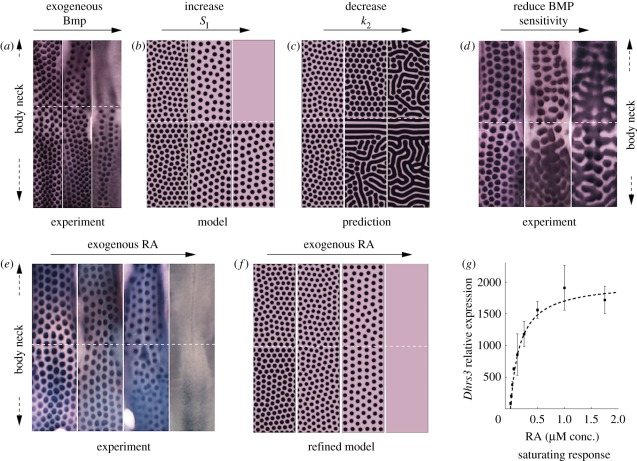
 to body
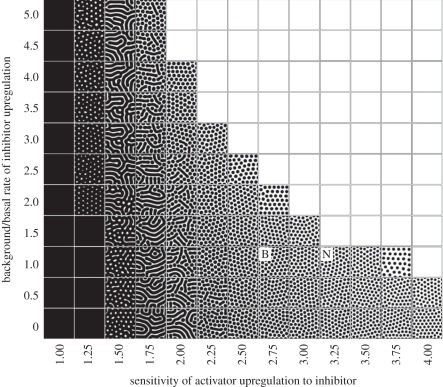
to body  to replicate the hypothesized variation in inhibitor sensitivity. Exogenous inhibitor is increased from left to right: control, SI = 1; SI = 1.5; SI = 2. (c) A model prediction showing the impact of uniformly reducing inhibitor sensitivity. From left to right: control,
to replicate the hypothesized variation in inhibitor sensitivity. Exogenous inhibitor is increased from left to right: control, SI = 1; SI = 1.5; SI = 2. (c) A model prediction showing the impact of uniformly reducing inhibitor sensitivity. From left to right: control,  . (d) An experimental reproduction shows the same qualitative behaviour. From left to right: control; +8 μM DM, +5 μM SB; +10 μM DM, +5 μM SB. DM, Dorsomorphin; SB, SB203580. (e) RA, which is normally only produced by neck skin, sensitizes the skin to the inhibitor. From left to right: control (no exogenous RA); +0.2 μM RA; +1.0 μM RA; +5.0 μM RA. (f) A refinement of the model in which k2 saturates according to equation (4.5) and recapitulates the experiment. We set kmax = 4, K = 5 and set varying RA levels for neck (RAn = 20 + RAe) and body (RAb = 10 + RAe) regions. From left to right: control, RAe = 0 ; RAe = 20; RAe = 100; RAe = 500. (g) Demonstration of saturability of RA signalling in cultured chicken skin. Each data point show the response of the skin to increasing concentrations of RA, measured through the relative expression of the RA response gene Dhrs3. Each point shows the mean expression and error bars, respectively. The RA dose–response curve was generated by quantitative PCR method using FastStart Universal SYBR Green Master Mix (Roche). The primer sequences used were: for Dhrs3, 5′-CTCTGCTGCCACCCAAAC-3′ and 5′-TGGTCTCCTTCAGGCATTTC-3; and for Gapdh, 5′-ATCTTTAACCACTGCTCCTTG-3′ and 5′-CATGCTGAGCCTATTCACTG-3′. The dashed line plots the saturating form of the function given by equation (4.5), with kmax = 2000, K = 0.175.
. (d) An experimental reproduction shows the same qualitative behaviour. From left to right: control; +8 μM DM, +5 μM SB; +10 μM DM, +5 μM SB. DM, Dorsomorphin; SB, SB203580. (e) RA, which is normally only produced by neck skin, sensitizes the skin to the inhibitor. From left to right: control (no exogenous RA); +0.2 μM RA; +1.0 μM RA; +5.0 μM RA. (f) A refinement of the model in which k2 saturates according to equation (4.5) and recapitulates the experiment. We set kmax = 4, K = 5 and set varying RA levels for neck (RAn = 20 + RAe) and body (RAb = 10 + RAe) regions. From left to right: control, RAe = 0 ; RAe = 20; RAe = 100; RAe = 500. (g) Demonstration of saturability of RA signalling in cultured chicken skin. Each data point show the response of the skin to increasing concentrations of RA, measured through the relative expression of the RA response gene Dhrs3. Each point shows the mean expression and error bars, respectively. The RA dose–response curve was generated by quantitative PCR method using FastStart Universal SYBR Green Master Mix (Roche). The primer sequences used were: for Dhrs3, 5′-CTCTGCTGCCACCCAAAC-3′ and 5′-TGGTCTCCTTCAGGCATTTC-3; and for Gapdh, 5′-ATCTTTAACCACTGCTCCTTG-3′ and 5′-CATGCTGAGCCTATTCACTG-3′. The dashed line plots the saturating form of the function given by equation (4.5), with kmax = 2000, K = 0.175.
References
-
- Klausmeier C. A. 1999. Regular and irregular patterns in semiarid vegetation. Science 284, 1826–182810.1126/science.284.5421.1826 (doi:10.1126/science.284.5421.1826) - DOI - DOI - PubMed
-
- Couzin I. D. 2009. Collective cognition in animal groups. Trends Cogn. Sci. 13, 36–4310.1016/j.tics.2008.10.002 (doi:10.1016/j.tics.2008.10.002) - DOI - DOI - PubMed
-
- Budrene E. O., Berg H. C. 1991. Complex patterns formed by motile cells of Escherichia coli. Nature 349, 630–63310.1038/349630a0 (doi:10.1038/349630a0) - DOI - DOI - PubMed
-
- Budrene E. O., Berg H. C. 1995. Dynamics of formation of symmetrical patterns by chemotactic bacteria. Nature 376, 49–5310.1038/376049a0 (doi:10.1038/376049a0) - DOI - DOI - PubMed
-
- Woodward D. E., Tyson R., Myerscough M. R., Murray J. D., Budrene E. O., Berg H. C. 1995. Spatio-temporal patterns generated by Salmonella typhimurium. Biophys. J. 68, 2181–218910.1016/S0006-3495(95)80400-5 (doi:10.1016/S0006-3495(95)80400-5) - DOI - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources