

Mitochondrial energetics is impaired in vivo in aged skeletal muscle
- PMID: 23919652
- PMCID: PMC4326861
- DOI: 10.1111/acel.12147
Mitochondrial energetics is impaired in vivo in aged skeletal muscle
Abstract
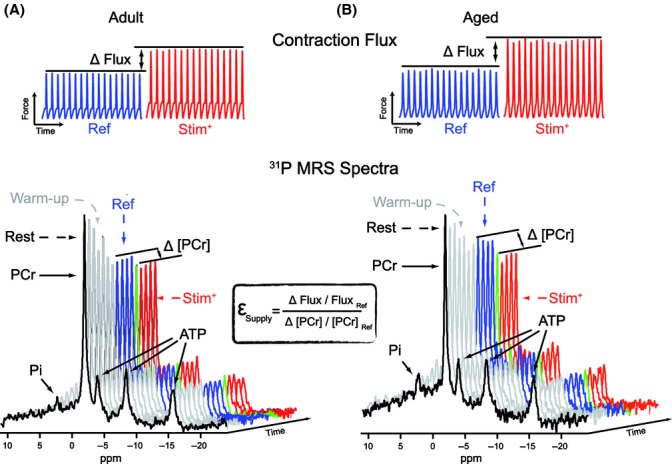
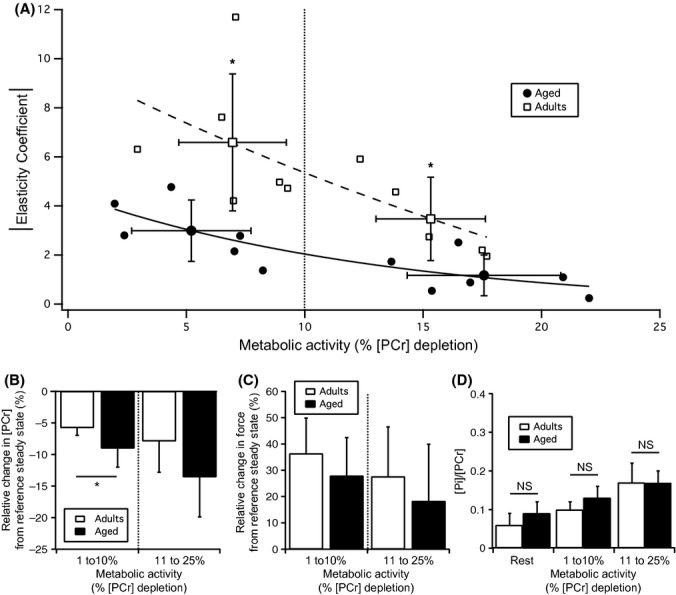
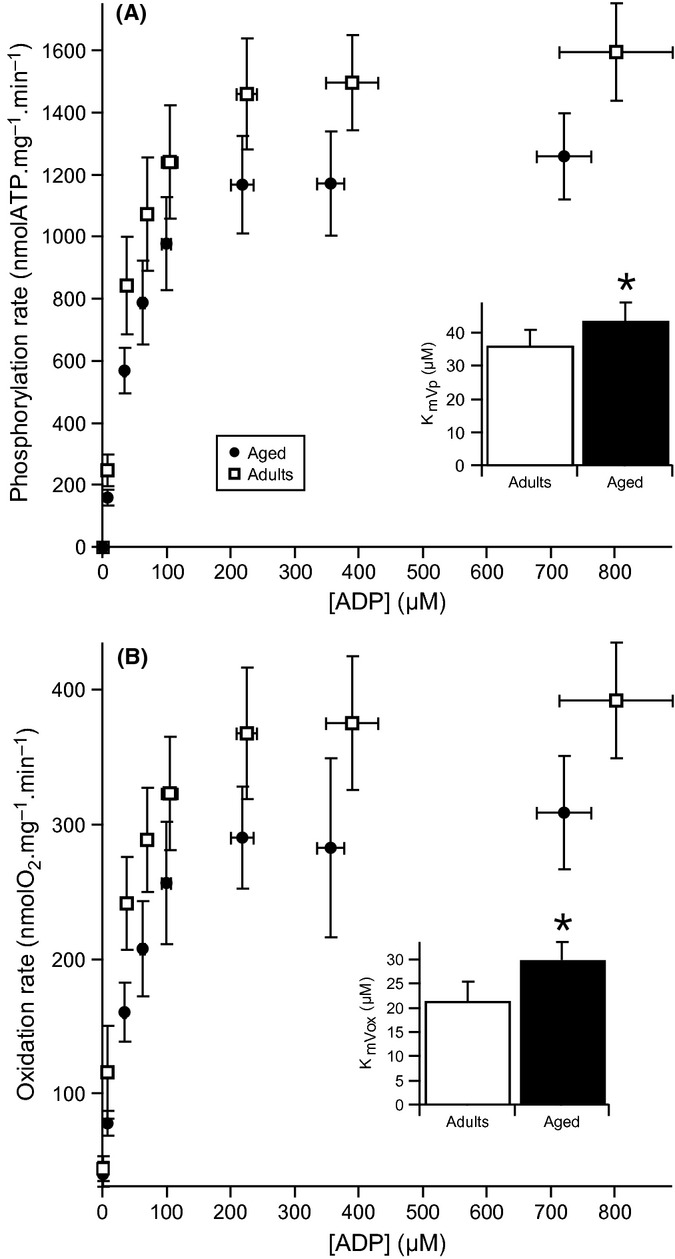
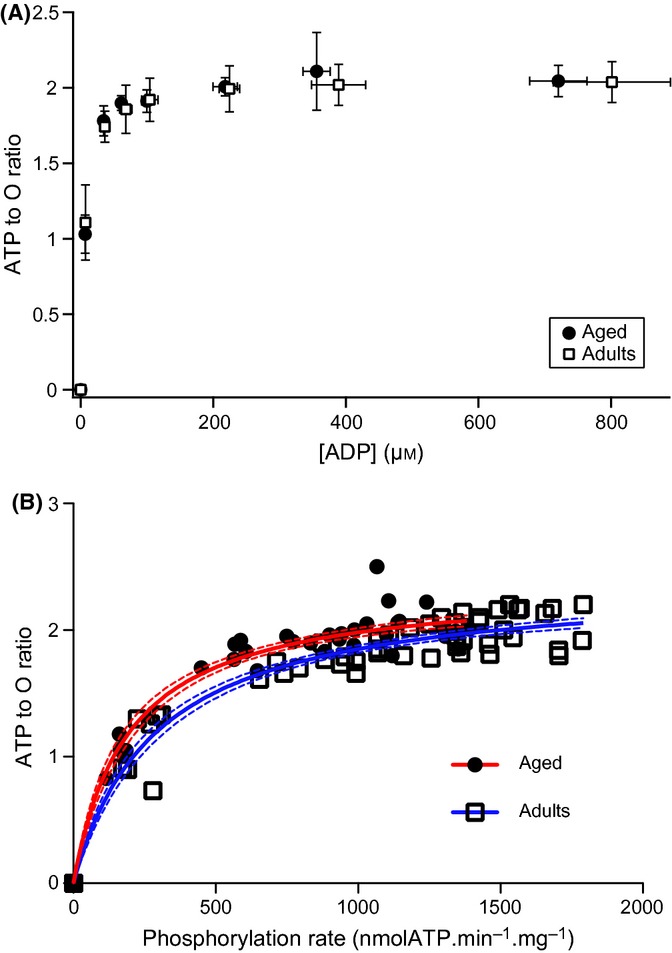
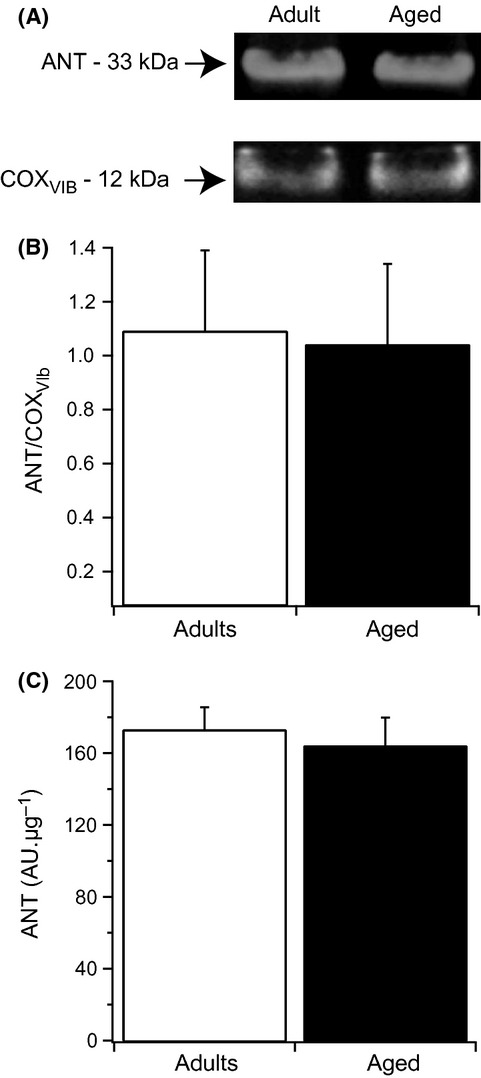
With aging, most skeletal muscles undergo a progressive loss of mass and strength, a process termed sarcopenia. Aging-related defects in mitochondrial energetics have been proposed to be causally involved in sarcopenia. However, changes in muscle mitochondrial oxidative phosphorylation with aging remain a highly controversial issue, creating a pressing need for integrative approaches to determine whether mitochondrial bioenergetics are impaired in aged skeletal muscle. To address this issue, mitochondrial bioenergetics was first investigated in vivo in the gastrocnemius muscle of adult (6 months) and aged (21 months) male Wistar rats by combining a modular control analysis approach with (31) P magnetic resonance spectroscopy measurements of energetic metabolites. Using this innovative approach, we revealed that the in vivo responsiveness ('elasticity') of mitochondrial oxidative phosphorylation to contraction-induced increase in ATP demand is significantly reduced in aged skeletal muscle, a reduction especially pronounced under low contractile activities. In line with this in vivo aging-related defect in mitochondrial energetics, we found that the mitochondrial affinity for ADP is significantly decreased in mitochondria isolated from aged skeletal muscle. Collectively, the results of this study demonstrate that mitochondrial bioenergetics are effectively altered in vivo in aged skeletal muscle and provide a novel cellular basis for this phenomenon.
Keywords: adenosine nucleotide translocator; affinity for ADP; energetics; in vivo; mitochondria; skeletal muscle aging.
© 2013 the Anatomical Society and John Wiley & Sons Ltd.
Figures
References
-
- Arsac LM, Beuste C, Miraux S, Deschodt-Arsac V, Thiaudiere E, Franconi JM, Diolez PH. In vivo modular control analysis of energy metabolism in contracting skeletal muscle. Biochem. J. 2008;414:391–397. - PubMed
-
- Bastien C, Sanchez J. Phosphagens and glycogen content in skeletal muscle after treadmill training in young and old rats. Eur. J. Appl. Physiol. Occup. Physiol. 1984;52:291–295. - PubMed
-
- Brand MD. Top down metabolic control analysis. J. Theor. Biol. 1996;182:351–360. - PubMed
-
- Brand MD, Curtis RK. Simplifying metabolic complexity. Biochem. Soc. Trans. 2002;30:25–30. - PubMed
-
- Bratic I, Trifunovic A. Mitochondrial energy metabolism and ageing. Biochim. Biophys. Acta. 2010;1797:961–967. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
