

Acinetobacter baumannii strain M2 produces type IV pili which play a role in natural transformation and twitching motility but not surface-associated motility
- PMID: 23919995
- PMCID: PMC3735195
- DOI: 10.1128/mBio.00360-13
Acinetobacter baumannii strain M2 produces type IV pili which play a role in natural transformation and twitching motility but not surface-associated motility
Abstract
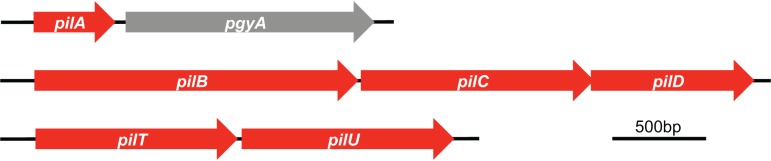
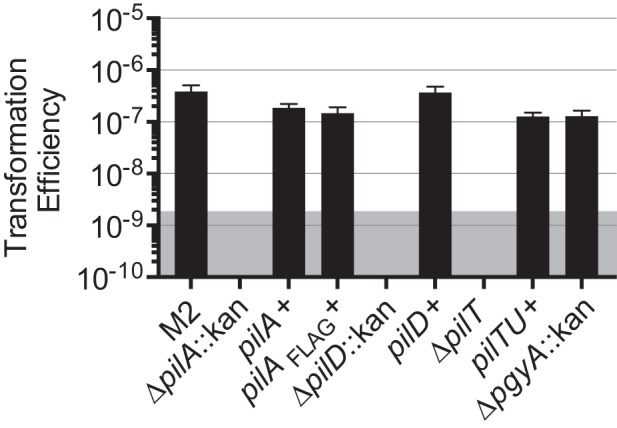
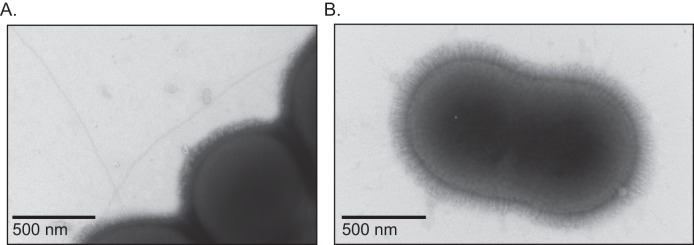
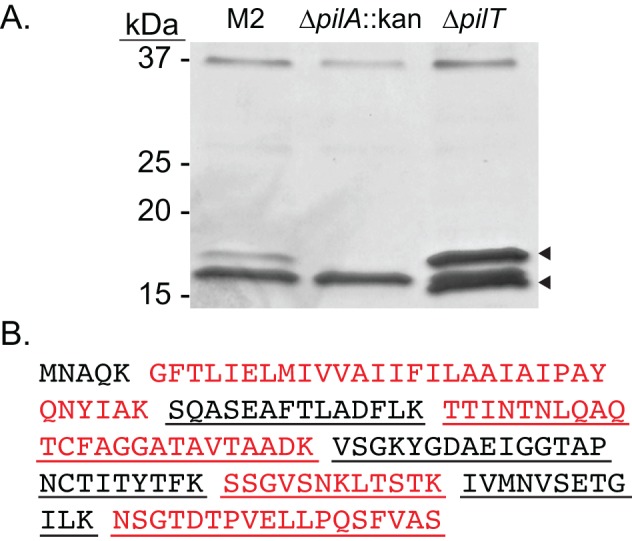
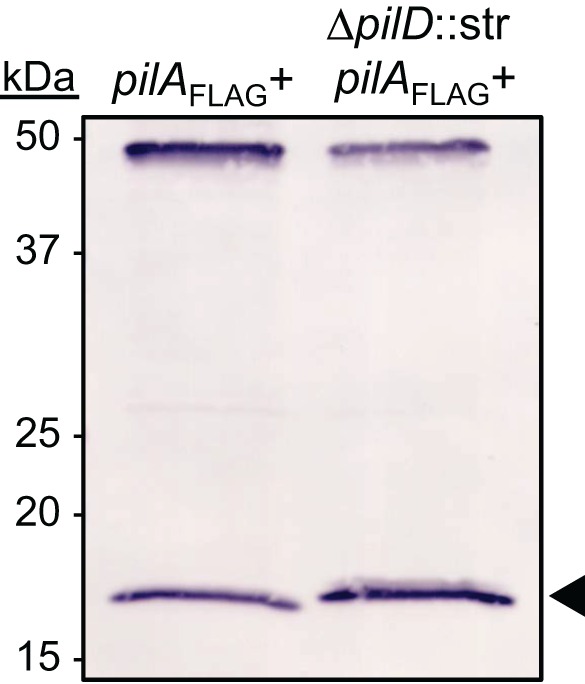
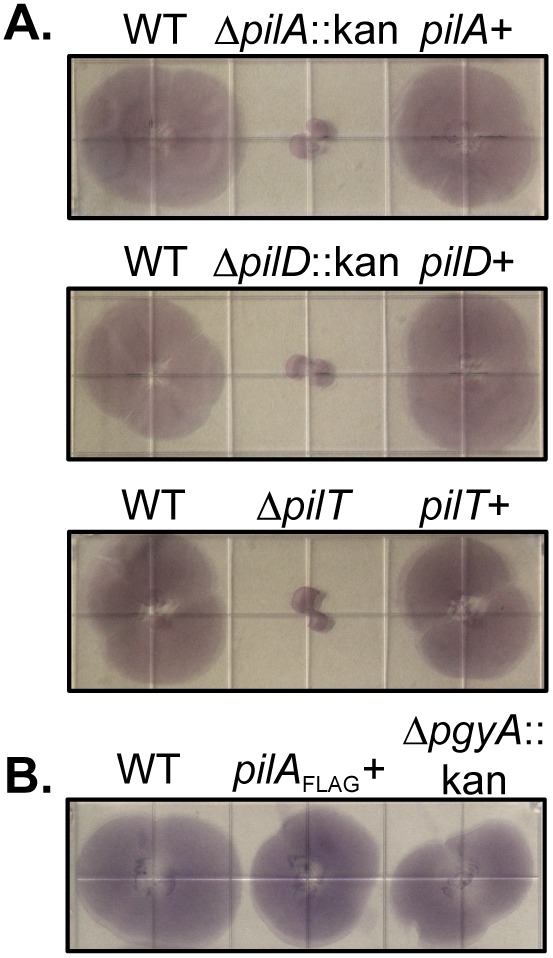
Acinetobacter baumannii is a Gram-negative, opportunistic pathogen. Recently, multiple A. baumannii genomes have been sequenced; these data have led to the identification of many genes predicted to encode proteins required for the biogenesis of type IV pili (TFP). However, there is no experimental evidence demonstrating that A. baumannii strains actually produce functional TFP. Here, we demonstrated that A. baumannii strain M2 is naturally transformable and capable of twitching motility, two classical TFP-associated phenotypes. Strains were constructed with mutations in pilA, pilD, and pilT, genes whose products have been well characterized in other systems. These mutants were no longer naturally transformable and did not exhibit twitching motility. These TFP-associated phenotypes were restored when these mutations were complemented. More PilA was detected on the surface of the pilT mutant than the parental strain, and TFP were visualized on the pilT mutant by transmission electron microscopy. Thus, A. baumannii produces functional TFP and utilizes TFP for both natural transformation and twitching motility. Several investigators have hypothesized that TFP might be responsible, in part, for the flagellum-independent surface-associated motility exhibited by many A. baumannii clinical isolates. We demonstrated that surface-associated motility was not dependent on the products of the pilA, pilD, and pilT genes and, by correlation, TFP. The identification of functional TFP in A. baumannii lays the foundation for future work determining the role of TFP in models of virulence that partially recapitulate human disease.
Importance: Several investigators have documented the presence of genes predicted to encode proteins required for the biogenesis of TFP in many A. baumannii genomes. Furthermore, some have speculated that TFP may play a role in the unique surface-associated motility phenotype exhibited by many A. baumannii clinical isolates, yet there has been no experimental evidence to prove this. Unfortunately, progress in understanding the biology and virulence of A. baumannii has been slowed by the difficulty of constructing and complementing mutations in this species. Strain M2, a recently characterized clinical isolate, is amenable to genetic manipulation. We have established a reproducible system for the generation of marked and/or unmarked mutations using a modified recombineering strategy as well as a genetic complementation system utilizing a modified mini-Tn7 element in strain M2. Using this strategy, we demonstrated that strain M2 produces TFP and that TFP are not required for surface-associated motility exhibited by strain M2.
Figures

References
-
- Maragakis LL, Perl TM. 2008. Acinetobacter baumannii: epidemiology, antimicrobial resistance, and treatment options. Clin. Infect. Dis. 46:1254–1263 - PubMed
-
- Dijkshoorn L, Nemec A, Seifert H. 2007. An increasing threat in hospitals: multidrug-resistant Acinetobacter baumannii. Nat. Rev. Microbiol. 5:939–951 - PubMed
-
- Towner KJ. 2009. Acinetobacter: an old friend, but a new enemy. J. Hosp. Infect. 73:355–363 - PubMed
-
- Weber DJ, Rutala WA, Miller MB, Huslage K, Sickbert-Bennett E. 2010. Role of hospital surfaces in the transmission of emerging health care-associated pathogens: norovirus, Clostridium difficile, and Acinetobacter species. Am. J. Infect. Control 38:S25–S33 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
