

Actin monomers activate inverted formin 2 by competing with its autoinhibitory interaction
- PMID: 23921379
- PMCID: PMC3772231
- DOI: 10.1074/jbc.M113.472415
Actin monomers activate inverted formin 2 by competing with its autoinhibitory interaction
Abstract
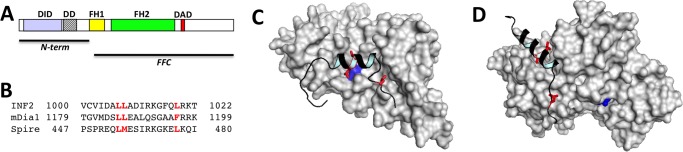
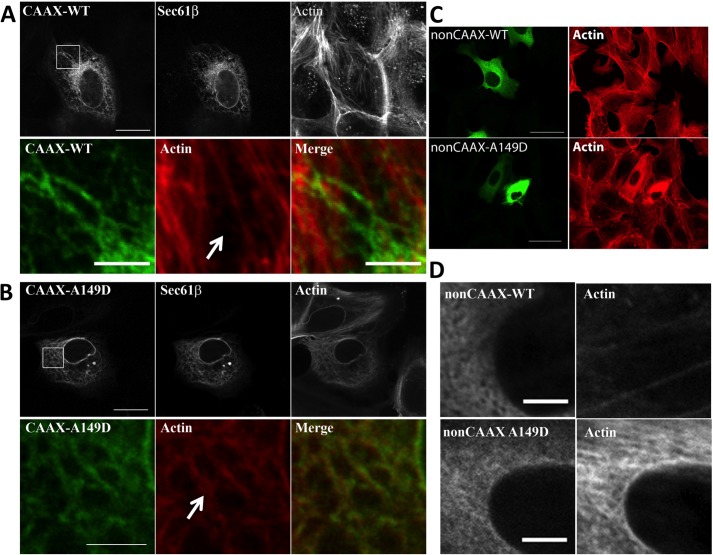
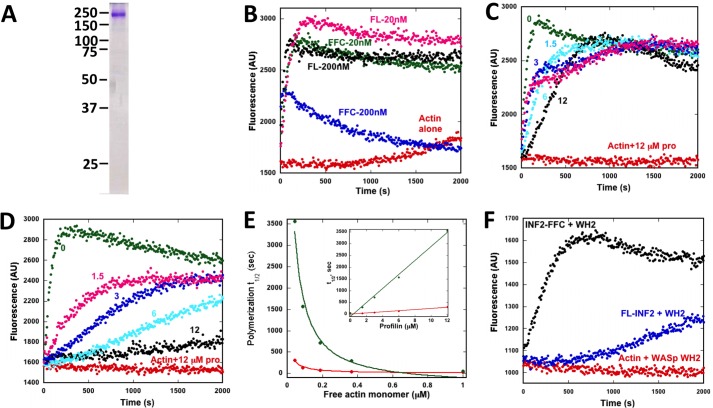
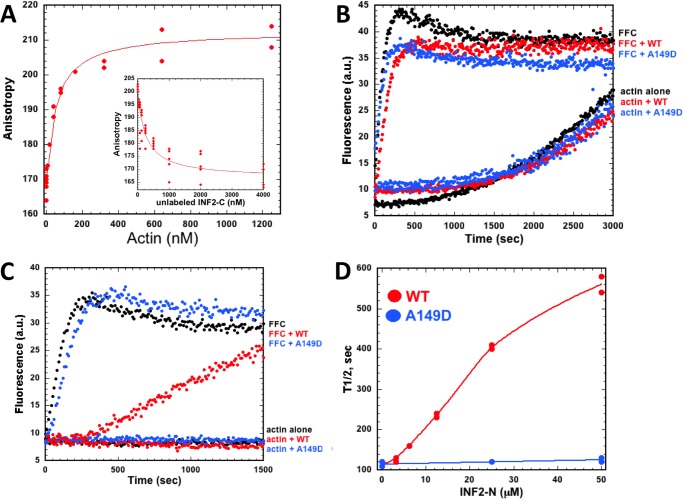
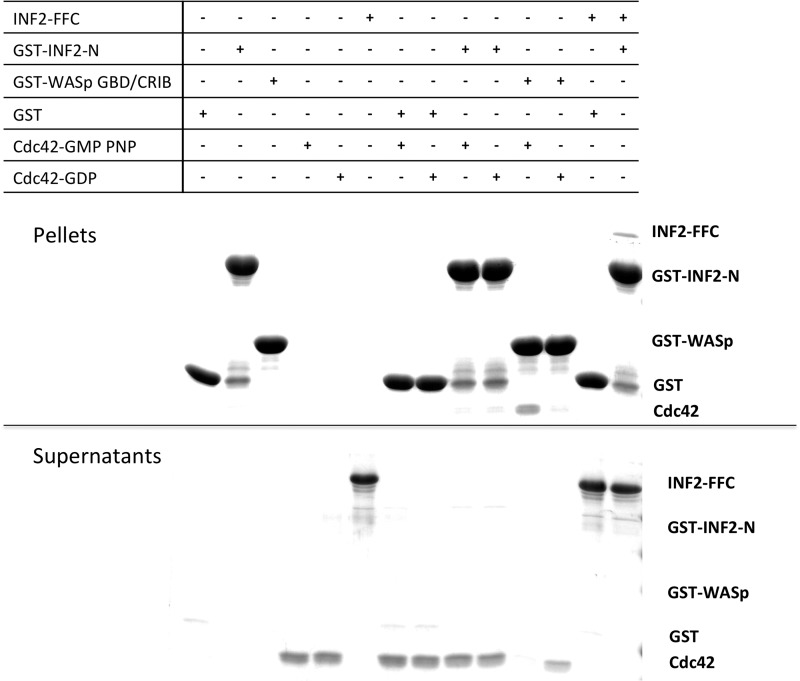
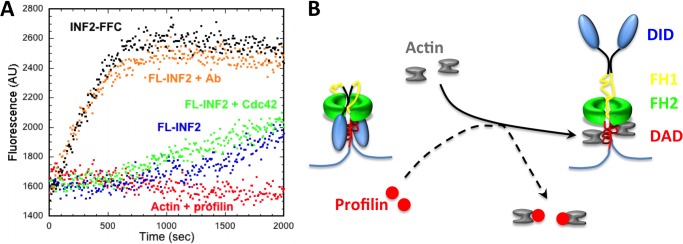
INF2 is an unusual formin protein in that it accelerates both actin polymerization and depolymerization, the latter through an actin filament-severing activity. Similar to other formins, INF2 possesses a dimeric formin homology 2 (FH2) domain that binds filament barbed ends and is critical for polymerization and depolymerization activities. In addition, INF2 binds actin monomers through its diaphanous autoregulatory domain (DAD) that resembles a Wiskott-Aldrich syndrome protein homology 2 (WH2) sequence C-terminal to the FH2 that participates in both polymerization and depolymerization. INF2-DAD is also predicted to participate in an autoinhibitory interaction with the N-terminal diaphanous inhibitory domain (DID). In this work, we show that actin monomer binding to the DAD of INF2 competes with the DID/DAD interaction, thereby activating actin polymerization. INF2 is autoinhibited in cells because mutation of a key DID residue results in constitutive INF2 activity. In contrast, purified full-length INF2 is constitutively active in biochemical actin polymerization assays containing only INF2 and actin monomers. Addition of proteins that compete with INF2-DAD for actin binding (profilin or the WH2 from Wiskott-Aldrich syndrome protein) decrease full-length INF2 activity while not significantly decreasing activity of an INF2 construct lacking the DID sequence. Profilin-mediated INF2 inhibition is relieved by an anti-N-terminal antibody for INF2 that blocks the DID/DAD interaction. These results suggest that free actin monomers can serve as INF2 activators by competing with the DID/DAD interaction. We also find that, in contrast to past results, the DID-containing N terminus of INF2 does not directly bind the Rho GTPase Cdc42.
Keywords: Actin; Cdc42; Endoplasmic Reticulum (ER); Formin; Rho.
Figures
Similar articles
-
INF2 Is a WASP homology 2 motif-containing formin that severs actin filaments and accelerates both polymerization and depolymerization.J Biol Chem. 2006 Sep 8;281(36):26754-67. doi: 10.1074/jbc.M604666200. Epub 2006 Jul 3. J Biol Chem. 2006. PMID: 16818491
-
INF2 is an endoplasmic reticulum-associated formin protein.J Cell Sci. 2009 May 1;122(Pt 9):1430-40. doi: 10.1242/jcs.040691. Epub 2009 Apr 14. J Cell Sci. 2009. PMID: 19366733 Free PMC article.
-
Mutations to the formin homology 2 domain of INF2 protein have unexpected effects on actin polymerization and severing.J Biol Chem. 2012 Oct 5;287(41):34234-45. doi: 10.1074/jbc.M112.365122. Epub 2012 Aug 9. J Biol Chem. 2012. PMID: 22879592 Free PMC article.
-
Formin proteins: a domain-based approach.Trends Biochem Sci. 2005 Jun;30(6):342-53. doi: 10.1016/j.tibs.2005.04.014. Trends Biochem Sci. 2005. PMID: 15950879 Review.
-
Role of formin INF2 in human diseases.Mol Biol Rep. 2022 Jan;49(1):735-746. doi: 10.1007/s11033-021-06869-x. Epub 2021 Oct 26. Mol Biol Rep. 2022. PMID: 34698992 Review.
Cited by
-
Proteomics analyses for the global proteins in the brain tissues of different human prion diseases.Mol Cell Proteomics. 2015 Apr;14(4):854-69. doi: 10.1074/mcp.M114.038018. Epub 2015 Jan 23. Mol Cell Proteomics. 2015. PMID: 25616867 Free PMC article.
-
Coordination of microtubule acetylation and the actin cytoskeleton by formins.Cell Mol Life Sci. 2018 Sep;75(17):3181-3191. doi: 10.1007/s00018-018-2855-3. Epub 2018 Jun 15. Cell Mol Life Sci. 2018. PMID: 29947928 Free PMC article. Review.
-
Calcium-mediated actin reset (CaAR) mediates acute cell adaptations.Elife. 2016 Dec 6;5:e19850. doi: 10.7554/eLife.19850. Elife. 2016. PMID: 27919320 Free PMC article.
-
Inverted formin 2 in focal adhesions promotes dorsal stress fiber and fibrillar adhesion formation to drive extracellular matrix assembly.Proc Natl Acad Sci U S A. 2015 May 12;112(19):E2447-56. doi: 10.1073/pnas.1505035112. Epub 2015 Apr 27. Proc Natl Acad Sci U S A. 2015. Retraction in: Proc Natl Acad Sci U S A. 2018 Mar 20;115(12):E2900. doi: 10.1073/pnas.1803125115. PMID: 25918420 Free PMC article. Retracted.
-
The actin-MRTF-SRF transcriptional circuit controls tubulin acetylation via α-TAT1 gene expression.J Cell Biol. 2018 Mar 5;217(3):929-944. doi: 10.1083/jcb.201702157. Epub 2018 Jan 10. J Cell Biol. 2018. PMID: 29321169 Free PMC article.
References
-
- Goode B. L., Eck M. J. (2007) Mechanism and function of formins in control of actin assembly. Annu. Rev. Biochem. 76, 593–627 - PubMed
-
- Li F., Higgs H. N. (2003) The mouse formin mDia1 is a potent actin nucleation factor regulated by autoinhibition. Curr. Biol. 13, 1335–1340 - PubMed
-
- Schulte A., Stolp B., Schönichen A., Pylypenko O., Rak A., Fackler O. T., Geyer M. (2008) The human formin FHOD1 contains a bipartite structure of FH3 and GTPase-binding domains required for activation. Structure 16, 1313–1323 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
