

Localized Fgf10 expression is not required for lung branching morphogenesis but prevents differentiation of epithelial progenitors
- PMID: 23924632
- PMCID: PMC3754473
- DOI: 10.1242/dev.096560
Localized Fgf10 expression is not required for lung branching morphogenesis but prevents differentiation of epithelial progenitors
Abstract
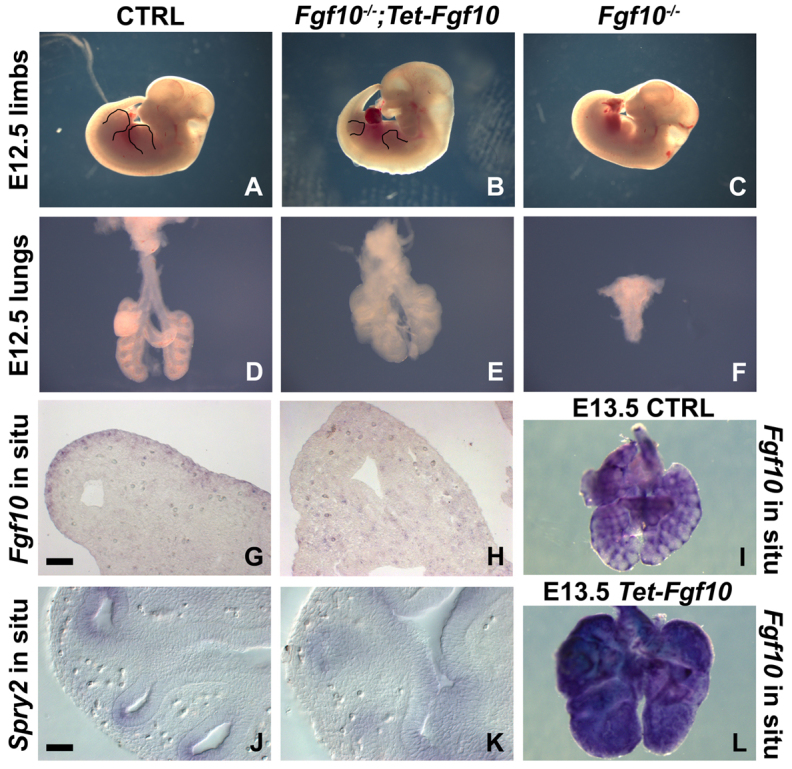
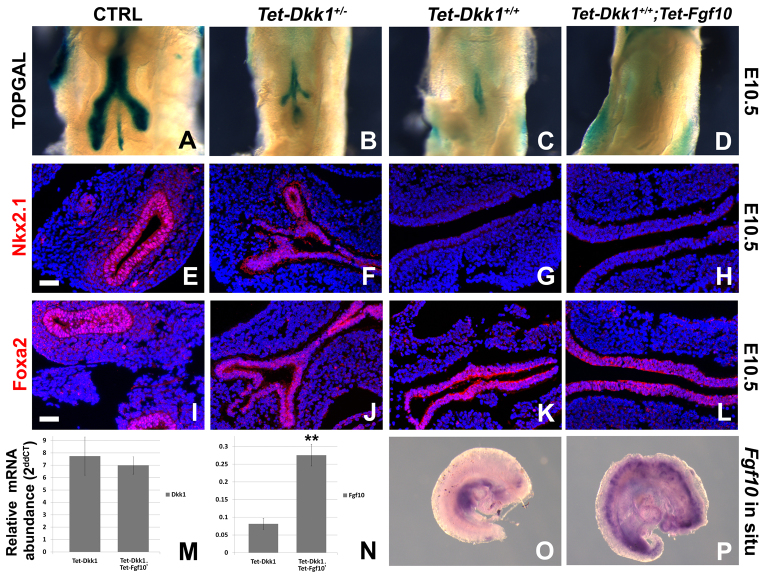
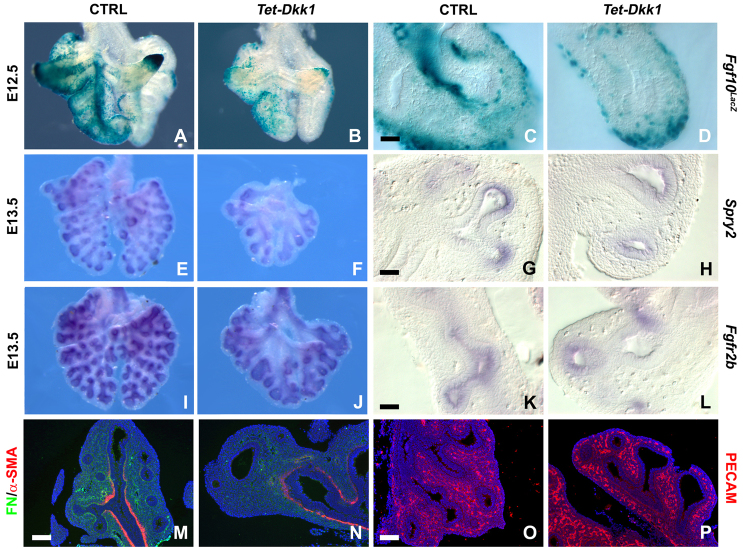
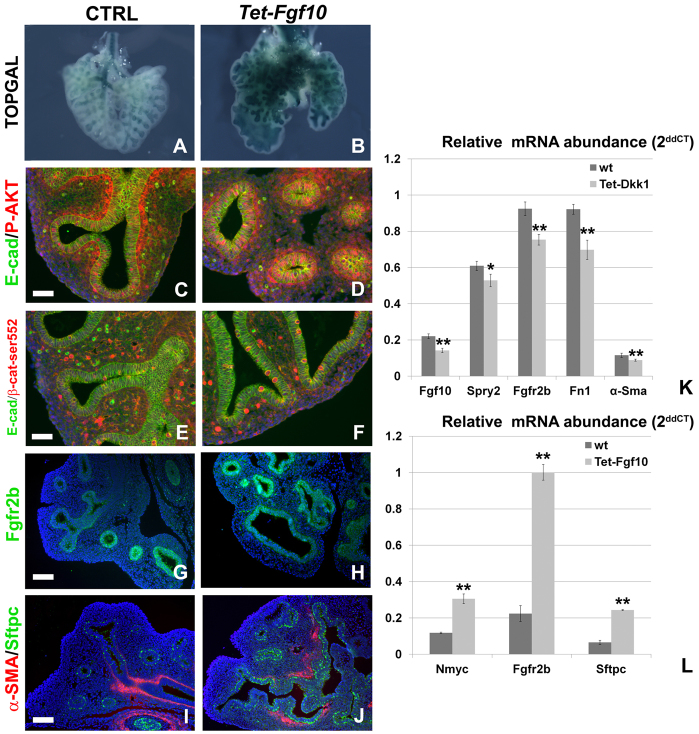
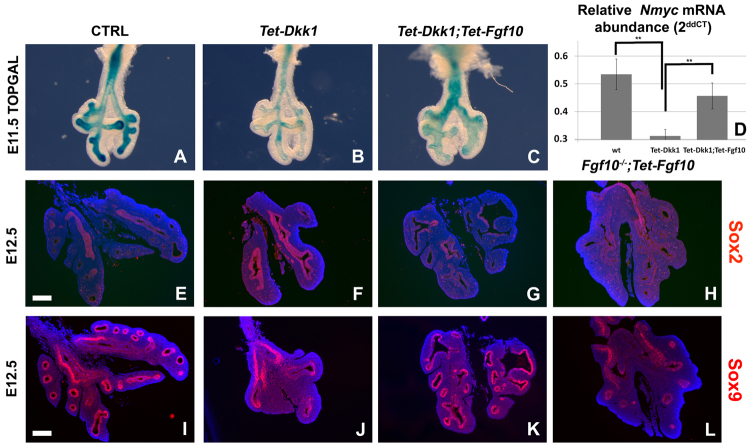
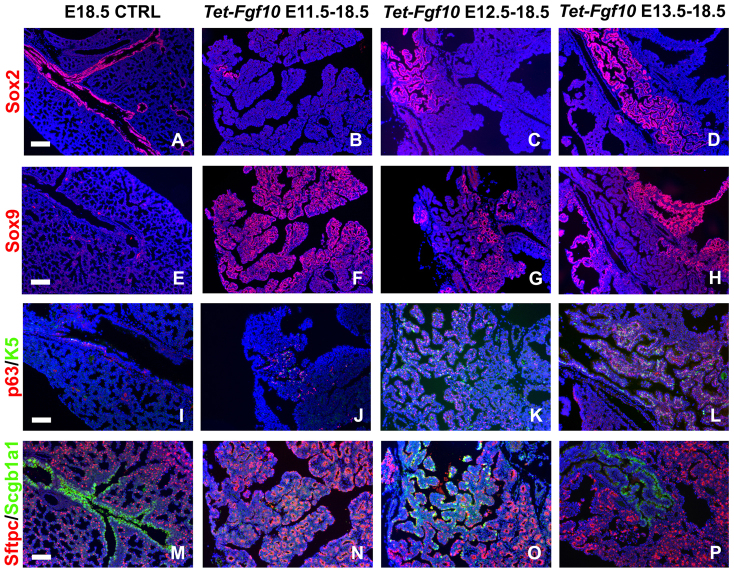
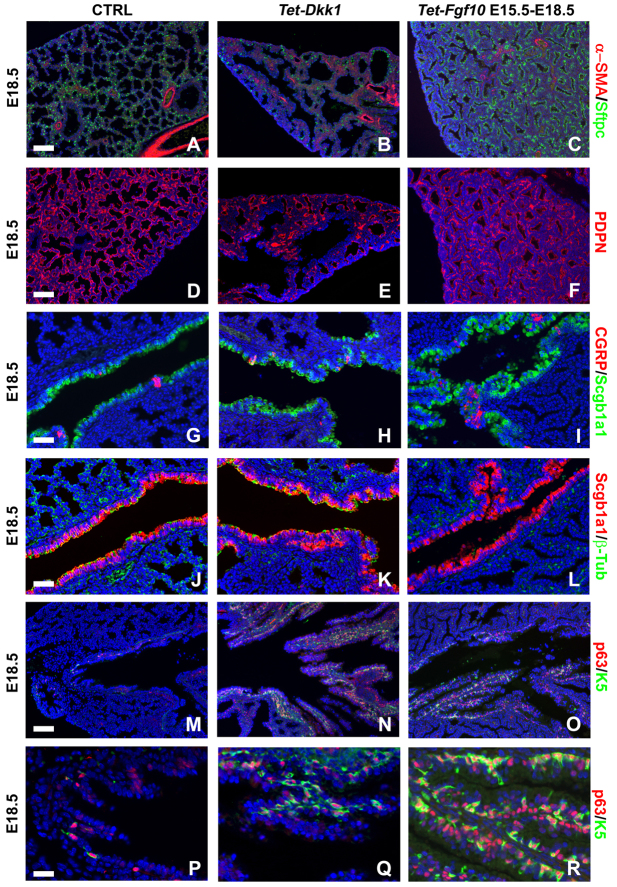
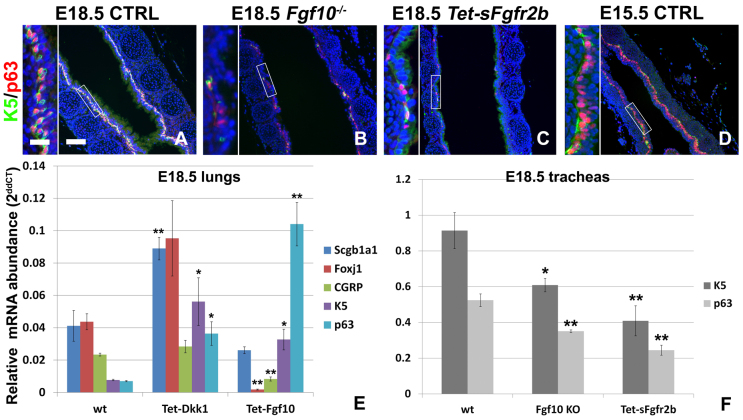
Localized Fgf10 expression in the distal mesenchyme adjacent to sites of lung bud formation has long been thought to drive stereotypic branching morphogenesis even though isolated lung epithelium branches in the presence of non-directional exogenous Fgf10 in Matrigel. Here, we show that lung agenesis in Fgf10 knockout mice can be rescued by ubiquitous overexpression of Fgf10, indicating that precisely localized Fgf10 expression is not required for lung branching morphogenesis in vivo. Fgf10 expression in the mesenchyme itself is regulated by Wnt signaling. Nevertheless, we found that during lung initiation simultaneous overexpression of Fgf10 is not sufficient to rescue the absence of primary lung field specification in embryos overexpressing Dkk1, a secreted inhibitor of Wnt signaling. However, after lung initiation, simultaneous overexpression of Fgf10 in lungs overexpressing Dkk1 is able to rescue defects in branching and proximal-distal differentiation. We also show that Fgf10 prevents the differentiation of distal epithelial progenitors into Sox2-expressing airway epithelial cells in part by activating epithelial β-catenin signaling, which negatively regulates Sox2 expression. As such, these findings support a model in which the main function of Fgf10 during lung development is to regulate proximal-distal differentiation. As the lung buds grow out, proximal epithelial cells become further and further displaced from the distal source of Fgf10 and differentiate into bronchial epithelial cells. Interestingly, our data presented here show that once epithelial cells are committed to the Sox2-positive airway epithelial cell fate, Fgf10 prevents ciliated cell differentiation and promotes basal cell differentiation.
Keywords: Basal cells; Branching; Dkk1; Fgf10; Lung development; Mouse; Wnt signaling.
Figures
References
-
- Ang S. L., Wierda A., Wong D., Stevens K. A., Cascio S., Rossant J., Zaret K. S. (1993). The formation and maintenance of the definitive endoderm lineage in the mouse: involvement of HNF3/forkhead proteins. Development 119, 1301–1315 - PubMed
-
- Bellusci S., Grindley J., Emoto H., Itoh N., Hogan B. L. (1997). Fibroblast growth factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 124, 4867–4878 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
