

Coordination of bacterial proteome with metabolism by cyclic AMP signalling
- PMID: 23925119
- PMCID: PMC4038431
- DOI: 10.1038/nature12446
Coordination of bacterial proteome with metabolism by cyclic AMP signalling
Abstract
The cyclic AMP (cAMP)-dependent catabolite repression effect in Escherichia coli is among the most intensely studied regulatory processes in biology. However, the physiological function(s) of cAMP signalling and its molecular triggers remain elusive. Here we use a quantitative physiological approach to show that cAMP signalling tightly coordinates the expression of catabolic proteins with biosynthetic and ribosomal proteins, in accordance with the cellular metabolic needs during exponential growth. The expression of carbon catabolic genes increased linearly with decreasing growth rates upon limitation of carbon influx, but decreased linearly with decreasing growth rate upon limitation of nitrogen or sulphur influx. In contrast, the expression of biosynthetic genes showed the opposite linear growth-rate dependence as the catabolic genes. A coarse-grained mathematical model provides a quantitative framework for understanding and predicting gene expression responses to catabolic and anabolic limitations. A scheme of integral feedback control featuring the inhibition of cAMP signalling by metabolic precursors is proposed and validated. These results reveal a key physiological role of cAMP-dependent catabolite repression: to ensure that proteomic resources are spent on distinct metabolic sectors as needed in different nutrient environments. Our findings underscore the power of quantitative physiology in unravelling the underlying functions of complex molecular signalling networks.
Figures
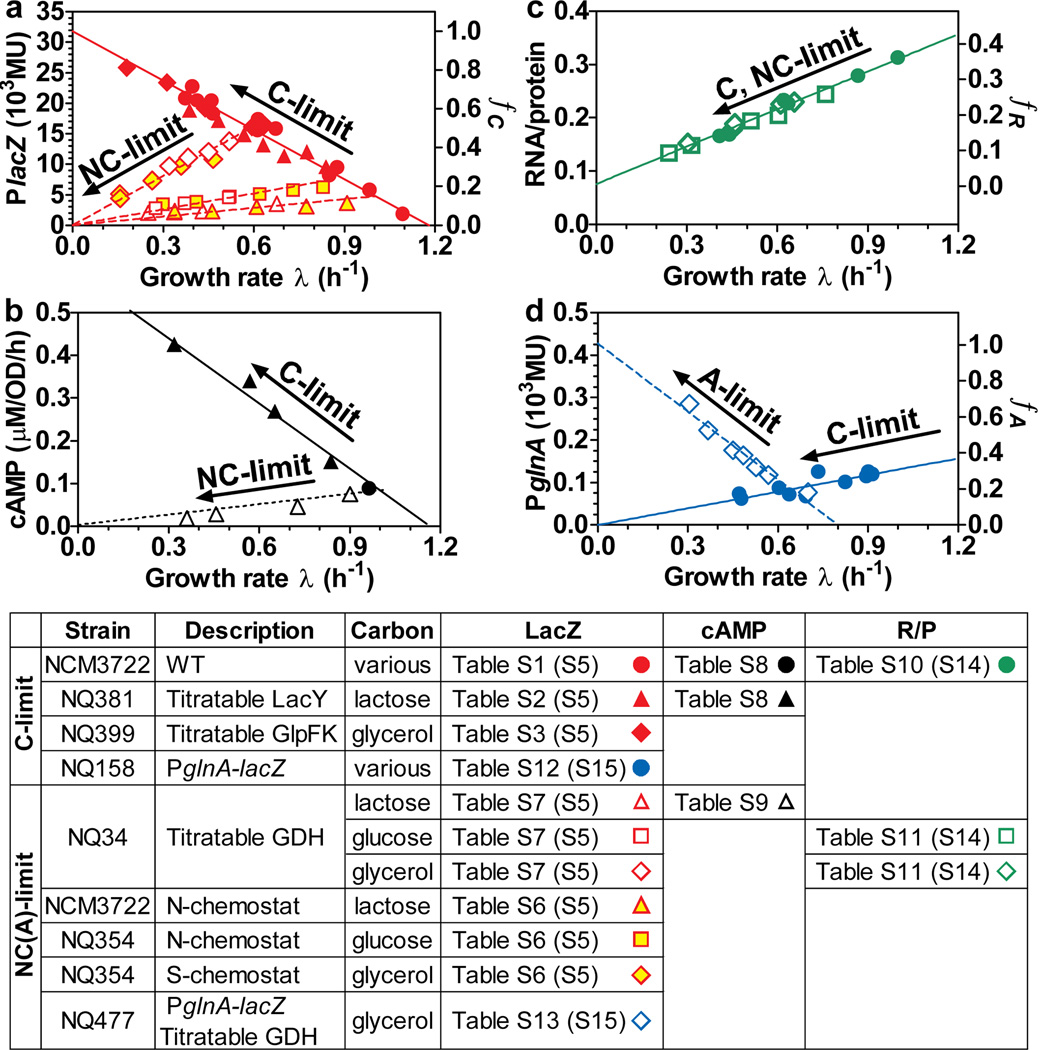
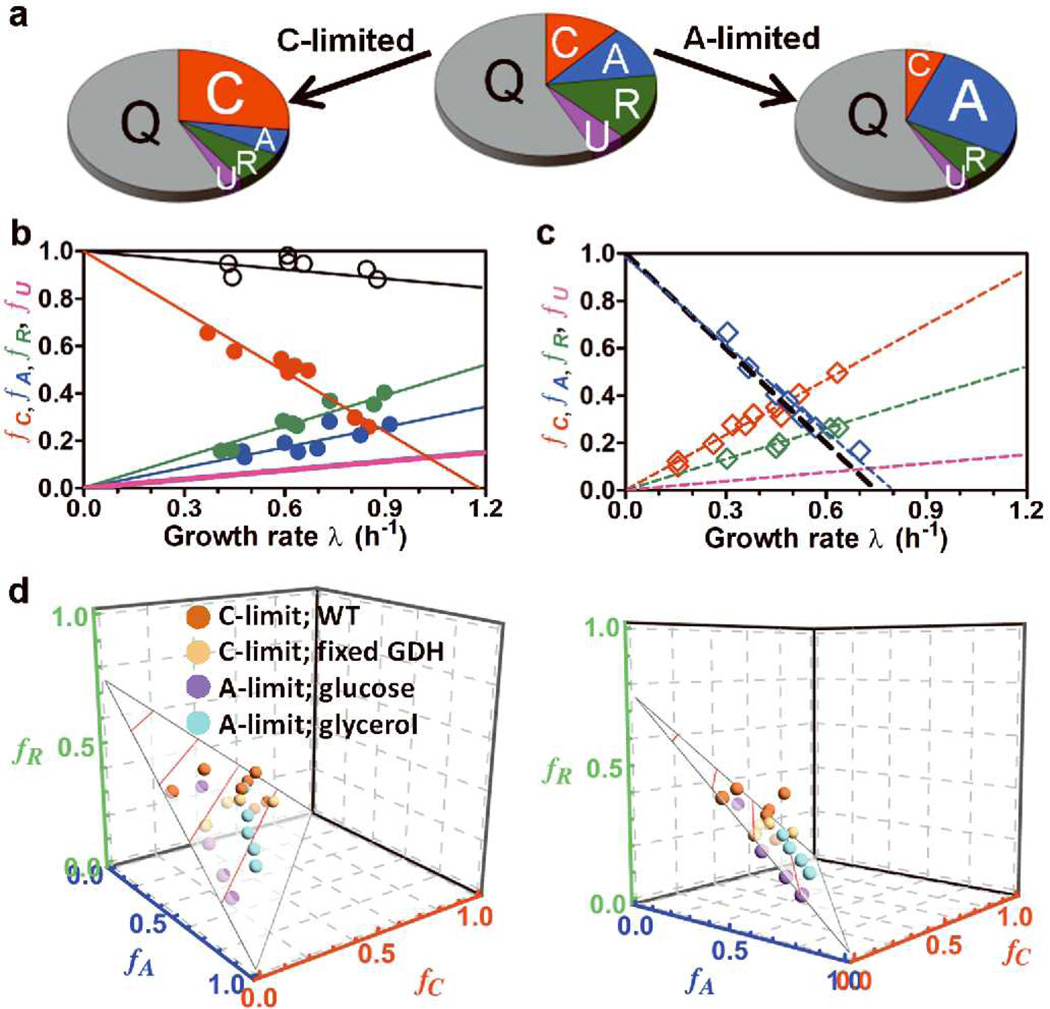


Comment in
-
Systems biology: metabolite turns master regulator.Nature. 2013 Aug 15;500(7462):283-4. doi: 10.1038/nature12544. Epub 2013 Aug 7. Nature. 2013. PMID: 23925111 Free PMC article.
References
-
- Laub MT, Goulian M. Specificity in two-component signal transduction pathways. Annu Rev Genet. 2007;41:121–145. - PubMed
-
- Potrykus K, Cashel M. (p)ppGpp: still magical? Annu Rev Microbiol. 2008;62:35–51. - PubMed
-
- Hengge R. Principles of c-di-GMP signalling in bacteria. Nature reviews. Microbiology. 2009;7:263–273. - PubMed
-
- Porter SL, Wadhams GH, Armitage JP. Signal processing in complex chemotaxis pathways. Nature reviews. Microbiology. 2011;9:153–165. - PubMed
-
- Brent R. Cell signaling: what is the signal and what information does it carry? FEBS Lett. 2009;583:4019–4024. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
