

The TFEB orthologue HLH-30 regulates autophagy and modulates longevity in Caenorhabditis elegans
- PMID: 23925298
- PMCID: PMC3866206
- DOI: 10.1038/ncomms3267
The TFEB orthologue HLH-30 regulates autophagy and modulates longevity in Caenorhabditis elegans
Abstract
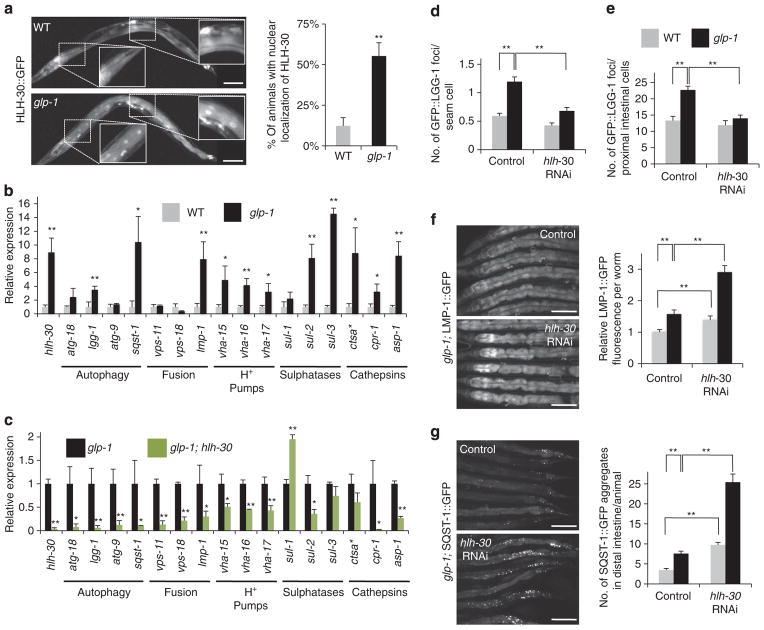
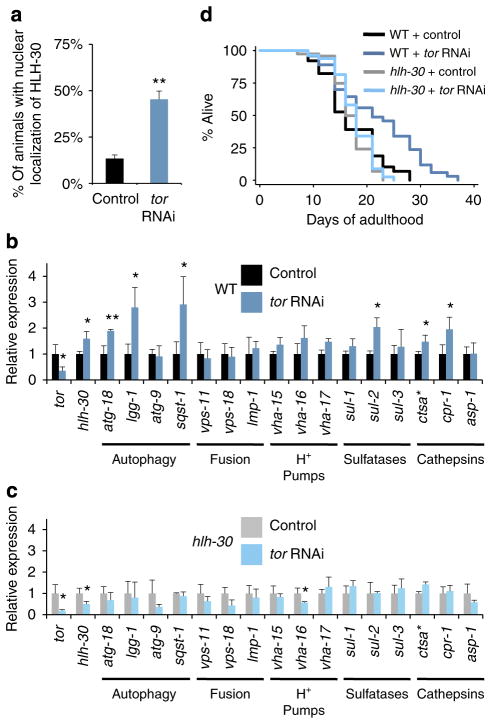
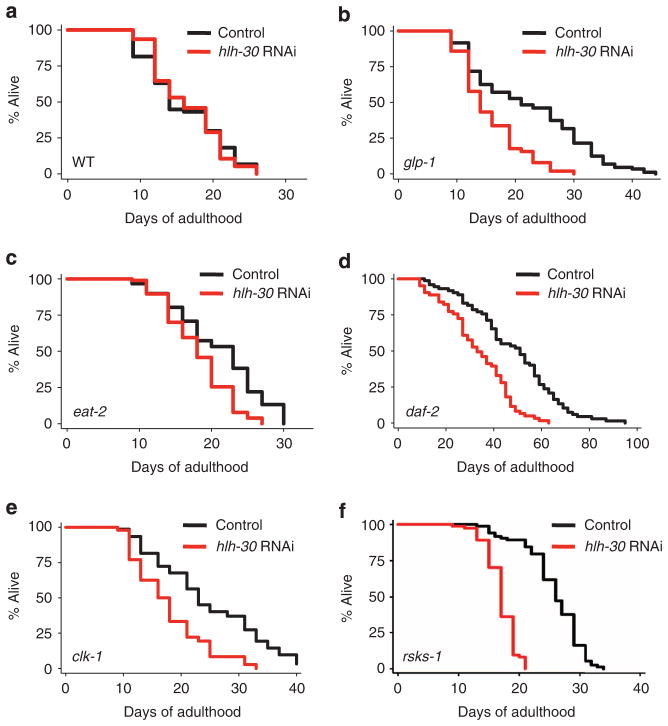
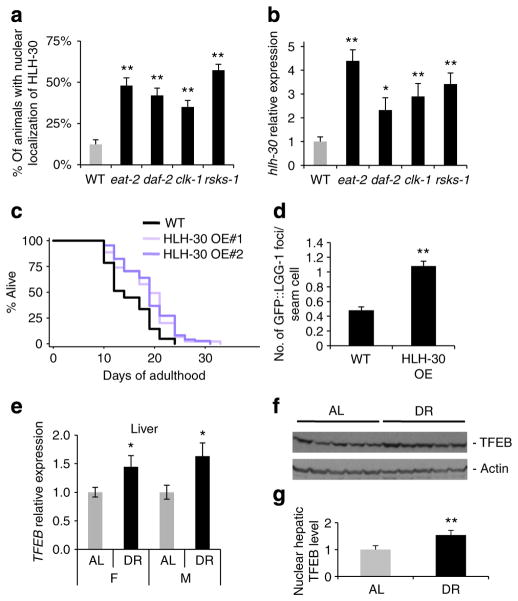
Autophagy is a cellular recycling process that has an important anti-aging role, but the underlying molecular mechanism is not well understood. The mammalian transcription factor EB (TFEB) was recently shown to regulate multiple genes in the autophagy process. Here we show that the predicted TFEB orthologue HLH-30 regulates autophagy in Caenorhabditis elegans and, in addition, has a key role in lifespan determination. We demonstrate that hlh-30 is essential for the extended lifespan of Caenorhabditis elegans in six mechanistically distinct longevity models, and overexpression of HLH-30 extends lifespan. Nuclear localization of HLH-30 is increased in all six Caenorhabditis elegans models and, notably, nuclear TFEB levels are augmented in the livers of mice subjected to dietary restriction, a known longevity-extending regimen. Collectively, our results demonstrate a conserved role for HLH-30 and TFEB in autophagy, and possibly longevity, and identify HLH-30 as a uniquely important transcription factor for lifespan modulation in Caenorhabditis elegans.
Conflict of interest statement
Figures
References
-
- Rubinsztein DC, Marino G, Kroemer G. Autophagy and aging. Cell. 2011;146:682–695. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
