

Neural mechanisms of speed perception: transparent motion
- PMID: 23926031
- PMCID: PMC3841929
- DOI: 10.1152/jn.00333.2013
Neural mechanisms of speed perception: transparent motion
Abstract
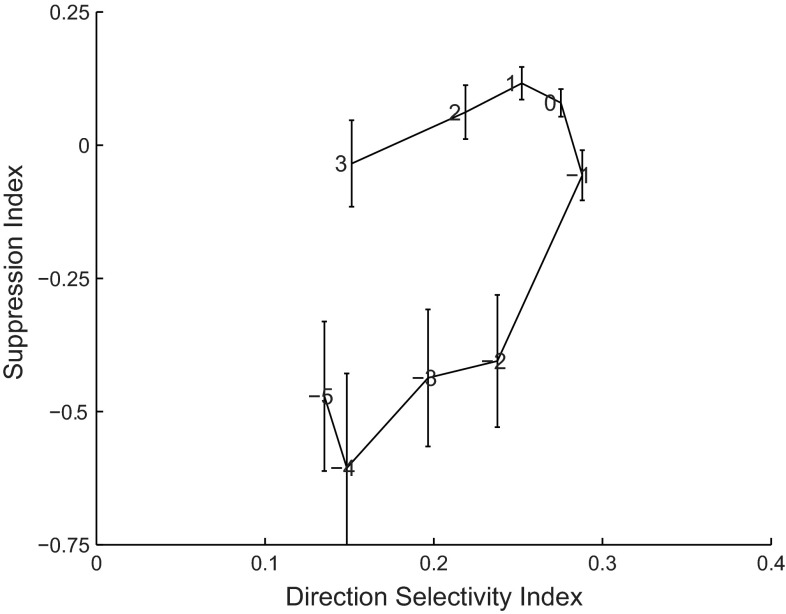
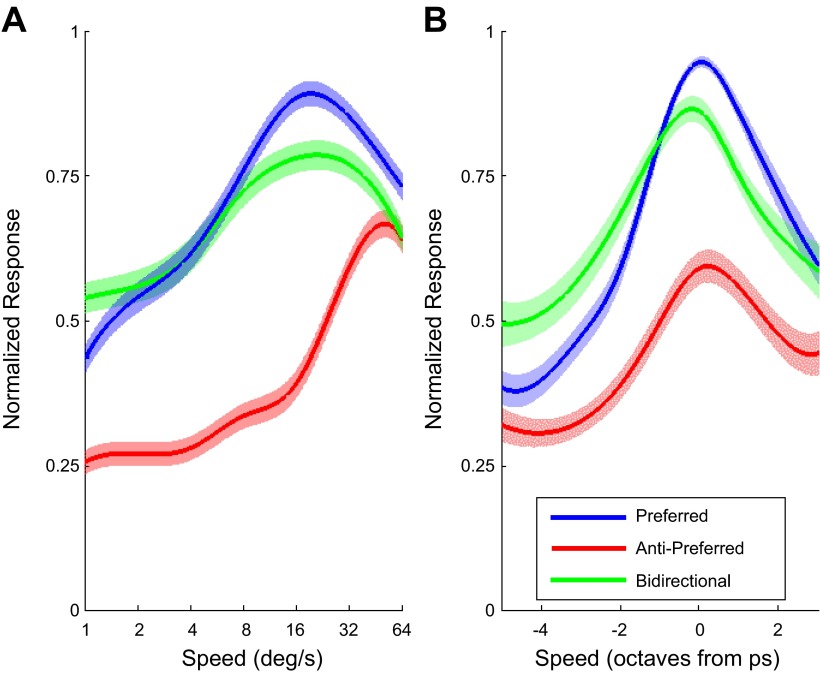
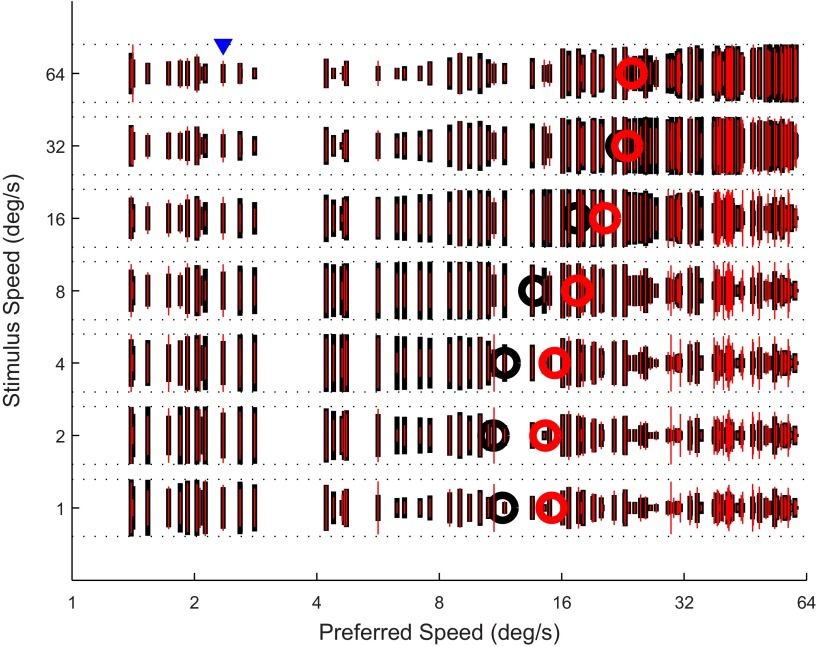
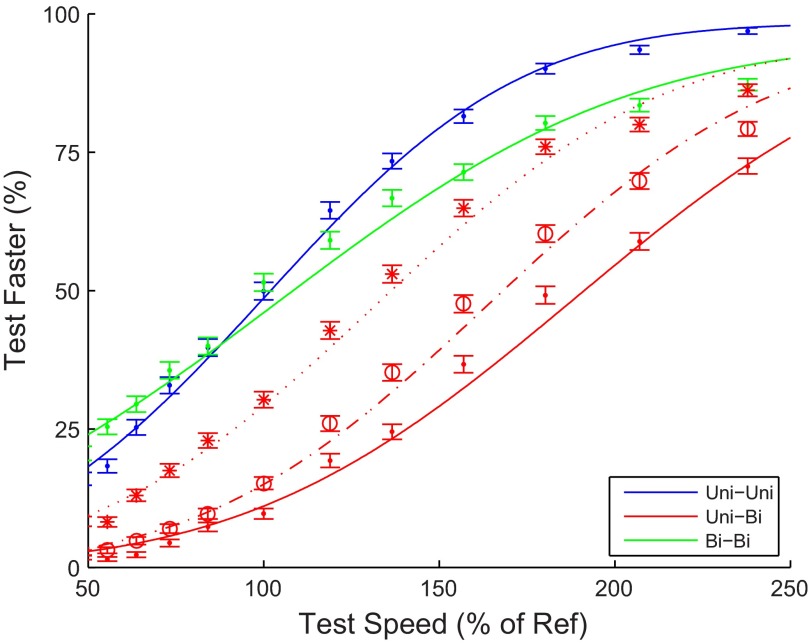
Visual motion on the macaque retina is processed by direction- and speed-selective neurons in extrastriate middle temporal cortex (MT). There is strong evidence for a link between the activity of these neurons and direction perception. However, there is conflicting evidence for a link between speed selectivity of MT neurons and speed perception. Here we study this relationship by using a strong perceptual illusion in speed perception: when two transparently superimposed dot patterns move in opposite directions, their apparent speed is much larger than the perceived speed of a single pattern moving at that physical speed. Moreover, the sensitivity for speed discrimination is reduced for such bidirectional patterns. We first confirmed these behavioral findings in human subjects and extended them to a monkey subject. Second, we determined speed tuning curves of MT neurons to bidirectional motion and compared these to speed tuning curves for unidirectional motion. Consistent with previous reports, the response to bidirectional motion was often reduced compared with unidirectional motion at the preferred speed. In addition, we found that tuning curves for bidirectional motion were shifted to lower preferred speeds. As a consequence, bidirectional motion of some speeds typically evoked larger responses than unidirectional motion. Third, we showed that these changes in neural responses could explain changes in speed perception with a simple labeled line decoder. These data provide new insight into the encoding of transparent motion patterns and provide support for the hypothesis that MT activity can be decoded for speed perception with a labeled line model.
Keywords: labeled line; macaque monkey; middle temporal area; motion perception; speed coding.
Figures
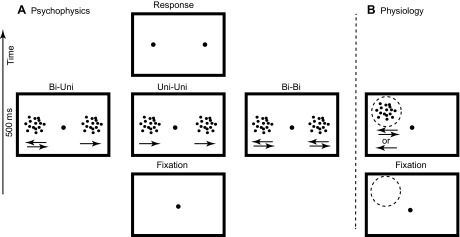
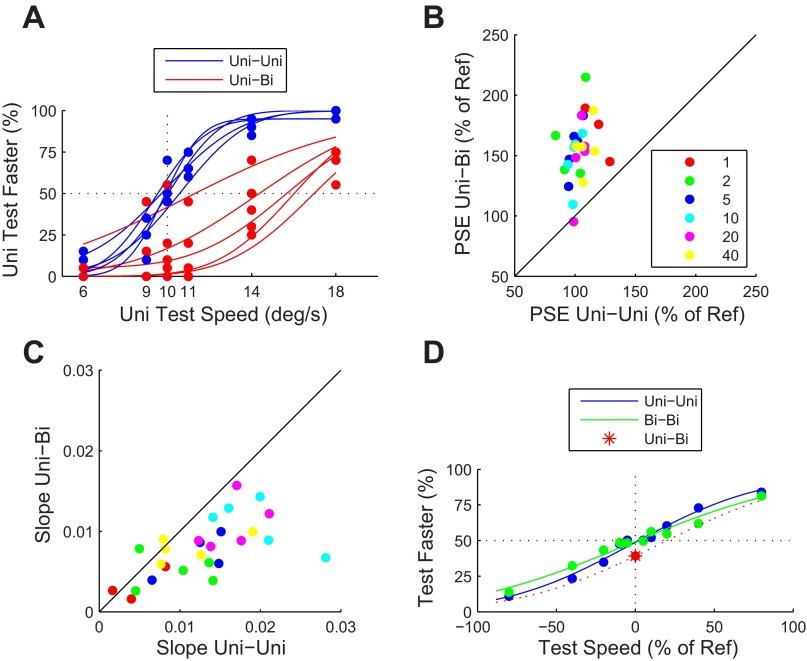
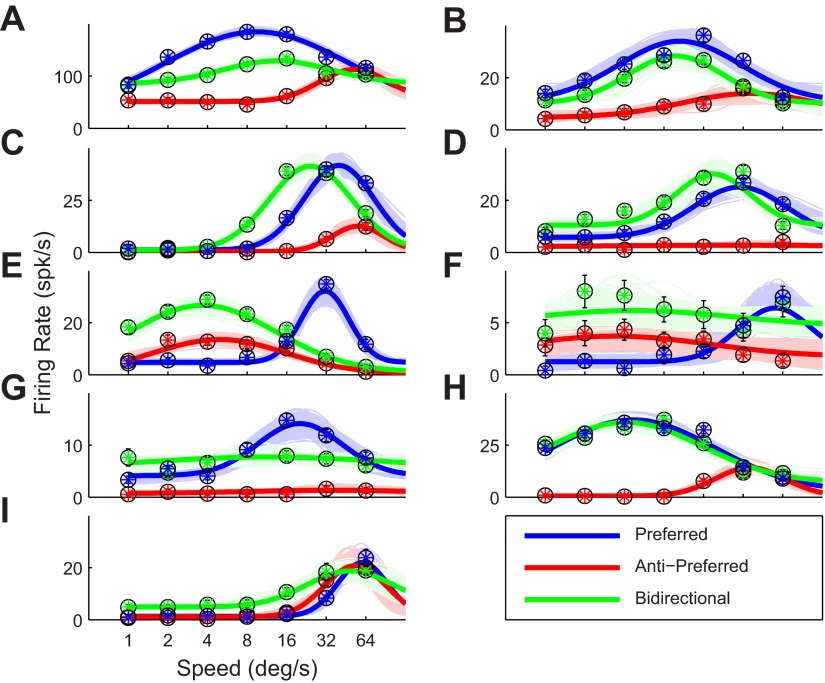
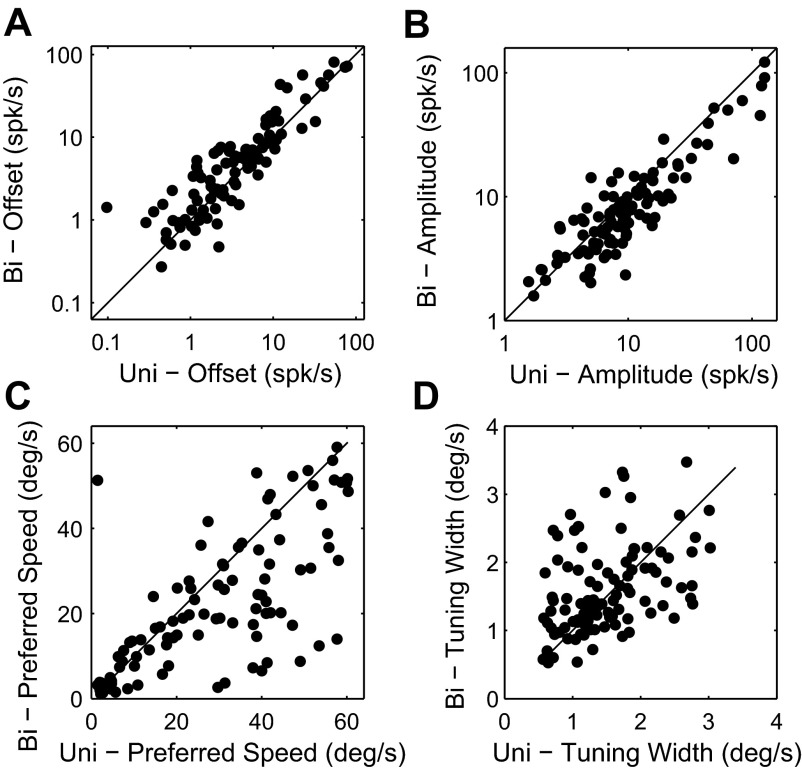
Similar articles
-
Misperceptions of speed are accounted for by the responses of neurons in macaque cortical area MT.J Neurophysiol. 2011 Mar;105(3):1199-211. doi: 10.1152/jn.00213.2010. Epub 2010 Dec 29. J Neurophysiol. 2011. PMID: 21191092 Free PMC article.
-
Distributed and Dynamic Neural Encoding of Multiple Motion Directions of Transparently Moving Stimuli in Cortical Area MT.J Neurosci. 2015 Dec 9;35(49):16180-98. doi: 10.1523/JNEUROSCI.2175-15.2015. J Neurosci. 2015. PMID: 26658869 Free PMC article.
-
Direction and orientation selectivity of neurons in visual area MT of the macaque.J Neurophysiol. 1984 Dec;52(6):1106-30. doi: 10.1152/jn.1984.52.6.1106. J Neurophysiol. 1984. PMID: 6520628
-
Motion selectivity in macaque visual cortex. III. Psychophysics and physiology of apparent motion.J Neurophysiol. 1986 Jun;55(6):1340-51. doi: 10.1152/jn.1986.55.6.1340. J Neurophysiol. 1986. PMID: 3734859
-
Motion selectivity in macaque visual cortex. I. Mechanisms of direction and speed selectivity in extrastriate area MT.J Neurophysiol. 1986 Jun;55(6):1308-27. doi: 10.1152/jn.1986.55.6.1308. J Neurophysiol. 1986. PMID: 3016210
Cited by
-
Criterion-free measurement of motion transparency perception at different speeds.J Vis. 2018 Apr 1;18(4):5. doi: 10.1167/18.4.5. J Vis. 2018. PMID: 29614154 Free PMC article.
-
Normalization of neuronal responses in cortical area MT across signal strengths and motion directions.J Neurophysiol. 2014 Sep 15;112(6):1291-306. doi: 10.1152/jn.00700.2013. Epub 2014 Jun 3. J Neurophysiol. 2014. PMID: 24899674 Free PMC article.
-
Evidence and Counterevidence in Motion Perception.Cereb Cortex. 2016 Dec;26(12):4602-4612. doi: 10.1093/cercor/bhv221. Epub 2015 Oct 3. Cereb Cortex. 2016. PMID: 26433972 Free PMC article.
-
Neural encoding of multiple motion speeds in visual cortical area MT.bioRxiv [Preprint]. 2023 Nov 22:2023.04.08.532456. doi: 10.1101/2023.04.08.532456. bioRxiv. 2023. PMID: 37070082 Free PMC article. Preprint.
-
Visual motion integration of bidirectional transparent motion in mouse opto-locomotor reflexes.Sci Rep. 2021 May 18;11(1):10490. doi: 10.1038/s41598-021-89974-y. Sci Rep. 2021. PMID: 34006985 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials