

The basolateral amygdala is critical for learning about neutral stimuli in the presence of danger, and the perirhinal cortex is critical in the absence of danger
- PMID: 23926265
- PMCID: PMC6619729
- DOI: 10.1523/JNEUROSCI.1998-13.2013
The basolateral amygdala is critical for learning about neutral stimuli in the presence of danger, and the perirhinal cortex is critical in the absence of danger
Abstract
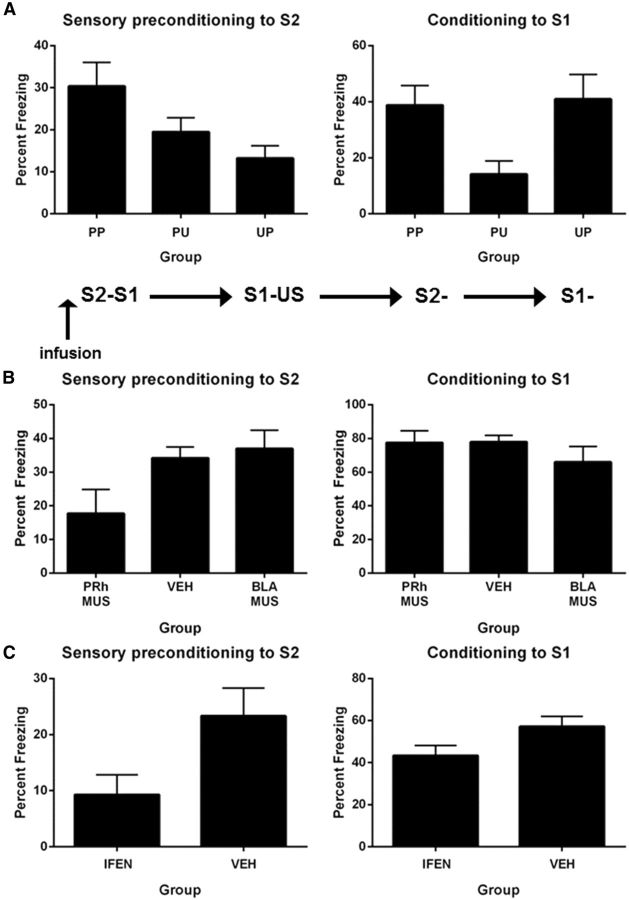
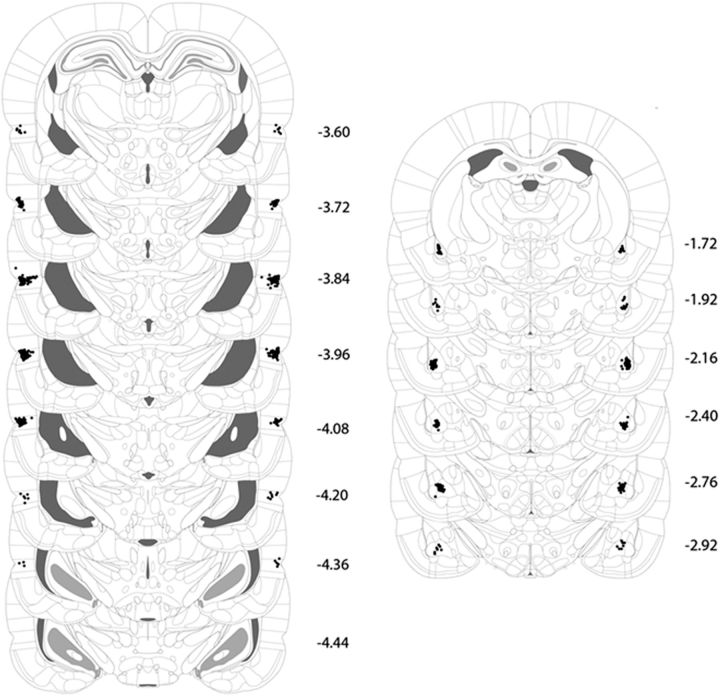
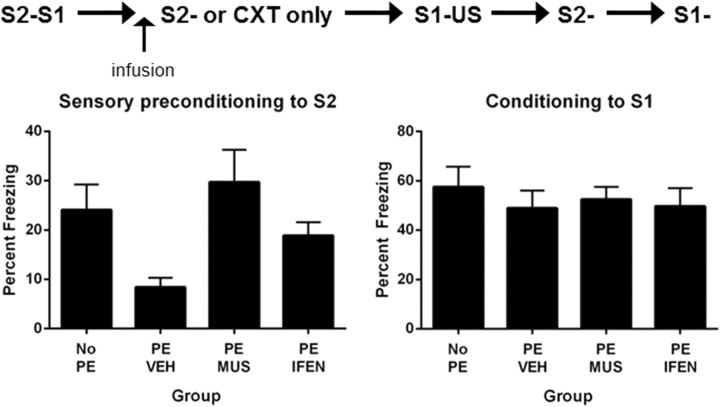
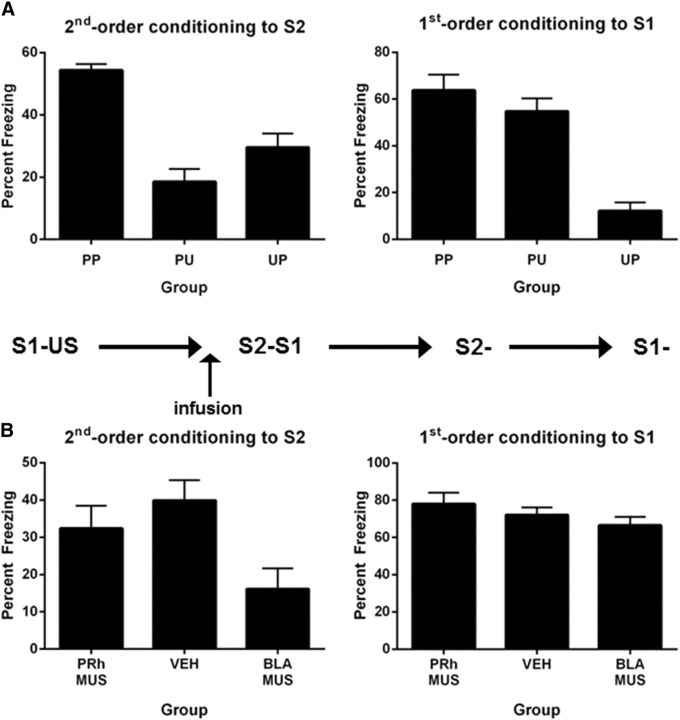
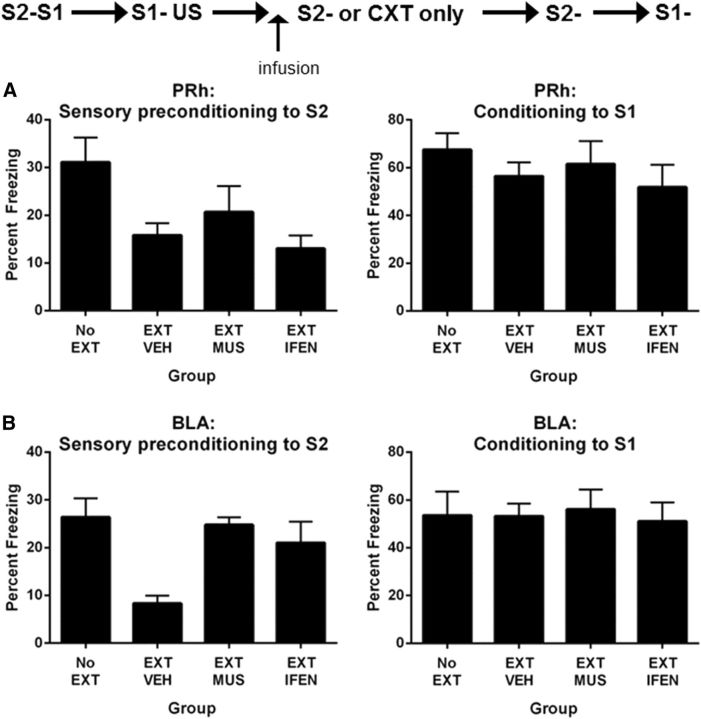
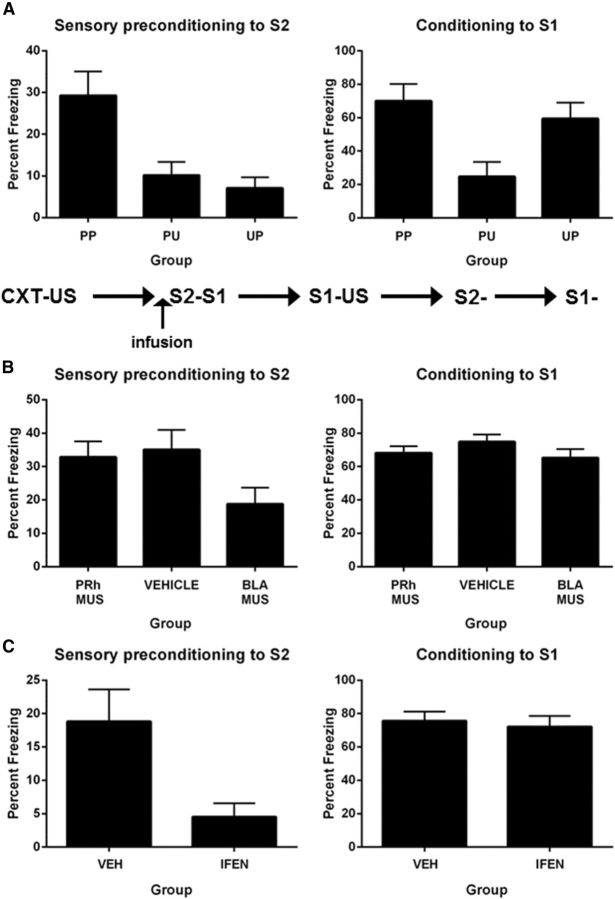
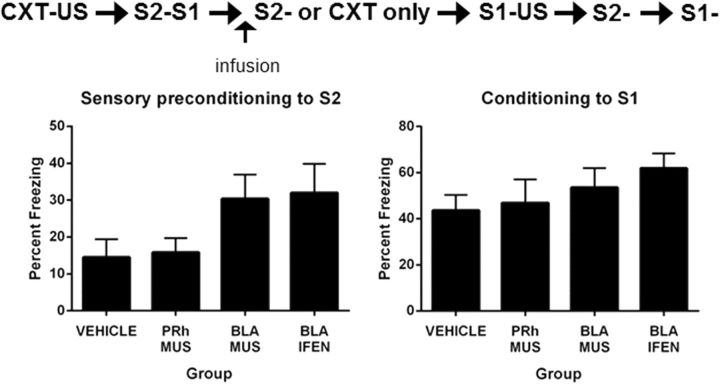
The perirhinal cortex (PRh) and basolateral amygdala (BLA) appear to mediate distinct aspects of learning and memory. Here, we used rats to investigate the involvement of the PRh and BLA in acquisition and extinction of associations between two different environmental stimuli (e.g., a tone and a light) in higher-order conditioning. When both stimuli were neutral, infusion of the GABAA, muscimol, or the NMDA receptor (NMDAR) antagonist ifenprodil into the PRh impaired associative formation. However, when one stimulus was neutral and the other was a learned danger signal, acquisition and extinction of the association between them was unaffected by manipulations targeting the PRh. Temporary inactivation of the BLA had the opposite effect: formation and extinction of an association between two stimuli was spared when both stimuli were neutral, but impaired when one stimulus was a learned danger signal. Subsequent experiments showed that the experience of fear per se shifts processing of an association between neutral stimuli from the PRh to the BLA. When training was conducted in a dangerous environment, formation and extinction of an association between neutral stimuli was impaired by BLA inactivation or NMDAR blockade in this region, but was unaffected by PRh inactivation. These double dissociations in the roles of the PRh and BLA in learning under different stimulus and environmental conditions imply that fear-induced activation of the amygdala changes how the brain processes sensory stimuli. Harmless stimuli are treated as potentially harmful, resulting in a shift from cortical to subcortical processing in the BLA.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources