

Human DNA Glycosylase NEIL1's Interactions with Downstream Repair Proteins Is Critical for Efficient Repair of Oxidized DNA Base Damage and Enhanced Cell Survival
- PMID: 23926464
- PMCID: PMC3733129
- DOI: 10.3390/biom2040564
Human DNA Glycosylase NEIL1's Interactions with Downstream Repair Proteins Is Critical for Efficient Repair of Oxidized DNA Base Damage and Enhanced Cell Survival
Abstract
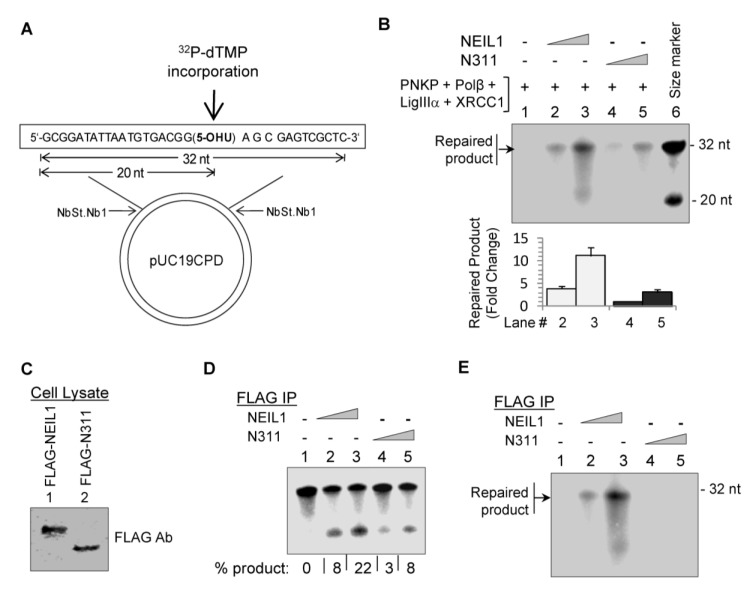
NEIL1 is unique among the oxidatively damaged base repair-initiating DNA glycosylases in the human genome due to its S phase-specific activation and ability to excise substrate base lesions from single-stranded DNA. We recently characterized NEIL1's specific binding to downstream canonical repair and non-canonical accessory proteins, all of which involve NEIL1's disordered C-terminal segment as the common interaction domain (CID). This domain is dispensable for NEIL1's base excision and abasic (AP) lyase activities, but is required for its interactions with other repair proteins. Here, we show that truncated NEIL1 lacking the CID is markedly deficient in initiating in vitro repair of 5-hydroxyuracil (an oxidative deamination product of C) in a plasmid substrate compared to the wild-type NEIL1, thus suggesting a critical role of CID in the coordination of overall repair. Furthermore, while NEIL1 downregulation significantly sensitized human embryonic kidney (HEK) 293 cells to reactive oxygen species (ROS), ectopic wild-type NEIL1, but not the truncated mutant, restored resistance to ROS. These results demonstrate that cell survival and NEIL1-dependent repair of oxidative DNA base damage require interactions among repair proteins, which could be explored as a cancer therapeutic target in order to increase the efficiency of chemo/radiation treatment.
Keywords: DNA glycosylase; NEIL1; base excision repair; common interaction domain; disordered structure; oxidative base damage and repair; protein-protein interaction; reactive oxygen species.
Figures




References
-
- Al-Tassan N., Chmiel N.H., Maynard J., Fleming N., Livingston A.L., Williams G.T., Hodges A.K., Davies D.R., David S.S., Sampson J.R., Cheadle J.P. Inherited variants of MYH associated with somatic G:C-->T:A mutations in colorectal tumors. Nat. Genet. 2002;30:227–232. doi: 10.1038/ng828. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous