

Dauer-specific dendrite arborization in C. elegans is regulated by KPC-1/Furin
- PMID: 23932402
- PMCID: PMC4671503
- DOI: 10.1016/j.cub.2013.06.058
Dauer-specific dendrite arborization in C. elegans is regulated by KPC-1/Furin
Abstract
Background: Dendrites often display remarkably complex and diverse morphologies that are influenced by developmental and environmental cues. Neuroplasticity in response to adverse environmental conditions entails both hypertrophy and resorption of dendrites. How dendrites rapidly alter morphology in response to unfavorable environmental conditions is unclear. The nematode Caenorhabditis elegans enters into a stress-resistant dauer larval stage in response to an adverse environment.
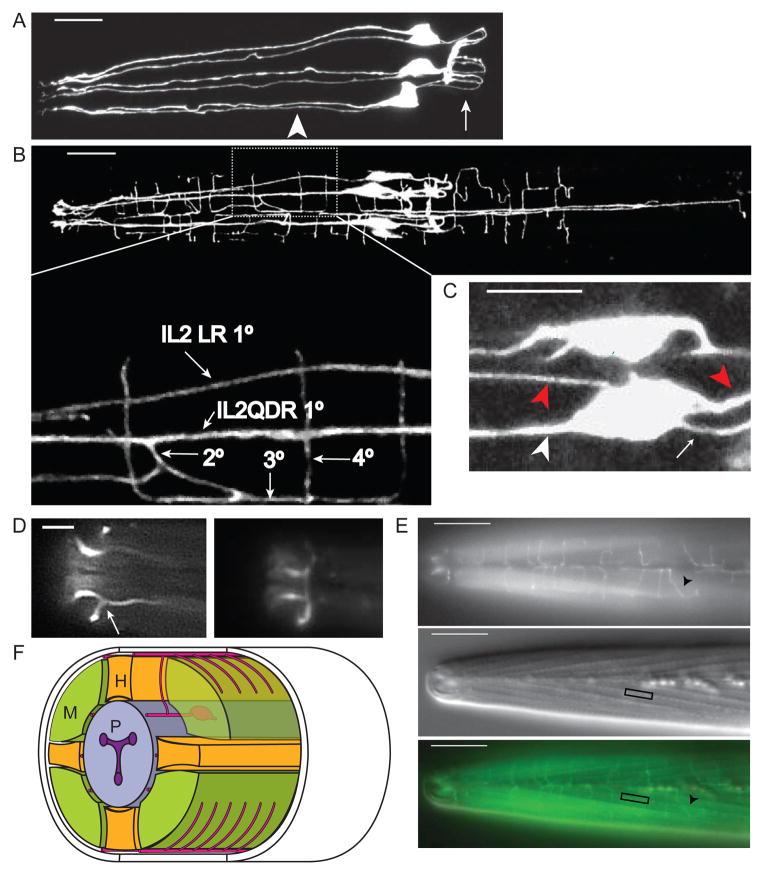
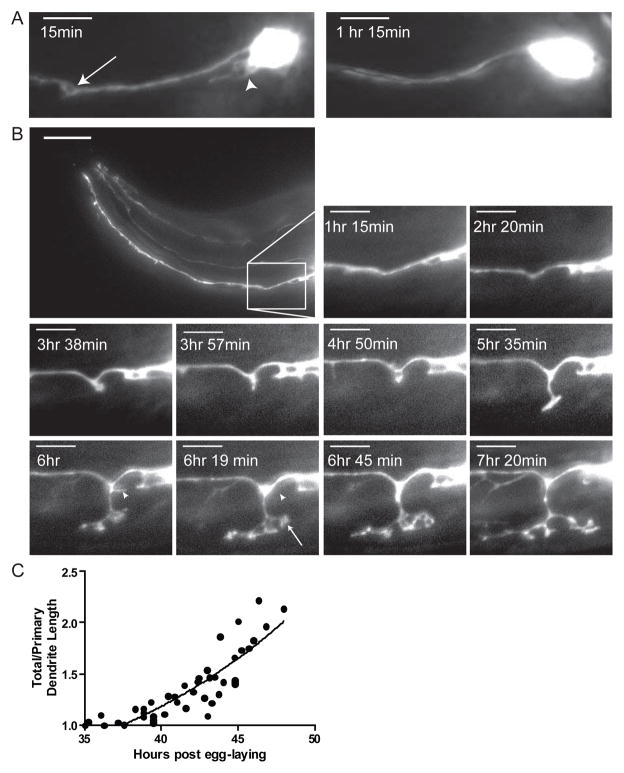
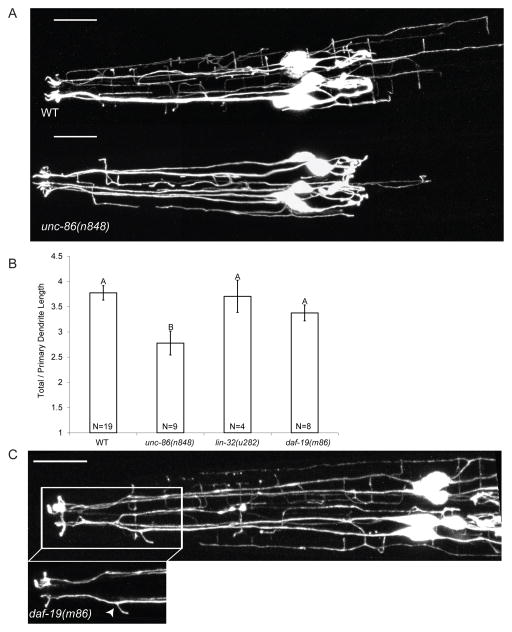
Results: Here we show that the IL2 bipolar sensory neurons undergo dendrite arborization and axon remodeling during dauer development. When dauer larvae are returned to favorable environmental conditions, animals resume reproductive development and IL2 dendritic branches retract, leaving behind remnant branches in postdauer L4 and adult animals. The C. elegans furin homolog KPC-1 is required for dauer IL2 dendritic arborization and dauer-specific nictation behavior. KPC-1 is also necessary for dendritic arborization of PVD and FLP sensory neurons. In mammals, furin is essential, ubiquitously expressed, and associated with numerous pathologies, including neurodegenerative diseases. While broadly expressed in C. elegans neurons and epithelia, KPC-1 acts cell autonomously in IL2 neurons to regulate dauer-specific dendritic arborization and nictation.
Conclusions: Neuroplasticity of the C. elegans IL2 sensory neurons provides a paradigm to study stress-induced and reversible dendritic branching, and the role of environmental and developmental cues in this process. The newly discovered role of KPC-1 in dendrite morphogenesis provides insight into the function of proprotein convertases in nervous system development.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures




Comment in
-
Dendrite plasticity: branching out for greener pastures.Curr Biol. 2013 Aug 19;23(16):R687-8. doi: 10.1016/j.cub.2013.07.036. Curr Biol. 2013. PMID: 23968924 Free PMC article.
References
-
- Riddle DL, Albert PS. In: Genetic and environmental regulation of dauer larvae developmentC. elegans. Riddle DL II, Blumenthal T, Meyer BJ, Priess JR, editors. Plainview, NY: Cold Spring Harbor Laboratory Press; 1998. pp. 739–768. - PubMed
-
- Cassada RC, Russell RL. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Dev Biol. 1975;2:326–342. - PubMed
-
- Lee H, Choi MK, Lee D, Kim HS, Hwang H, Kim H, Park S, Paik YK, Lee J. Nictation, a dispersal behavior of the nematode Caenorhabditis elegans, is regulated by IL2 neurons. Nat Neurosci. 2011;1:107–112. - PubMed
-
- Golden JW, Riddle DL. The Caenorhabditis elegans dauer larva: developmental effects of pheromone, food, and temperature. Dev Biol. 1984;2:368–378. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
