

Structures of human folate receptors reveal biological trafficking states and diversity in folate and antifolate recognition
- PMID: 23934049
- PMCID: PMC3780903
- DOI: 10.1073/pnas.1308827110
Structures of human folate receptors reveal biological trafficking states and diversity in folate and antifolate recognition
Abstract
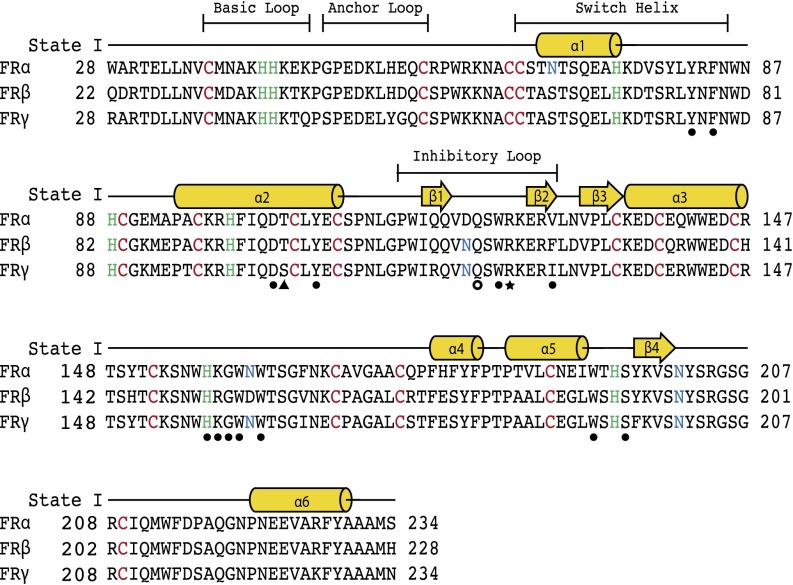
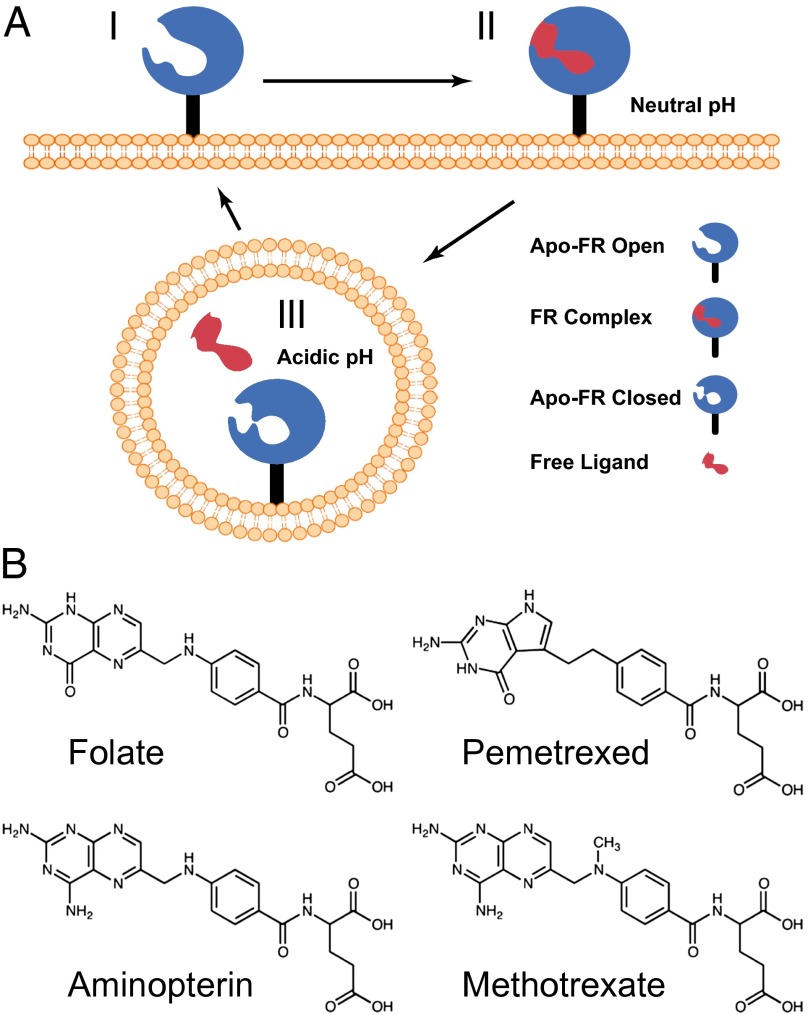
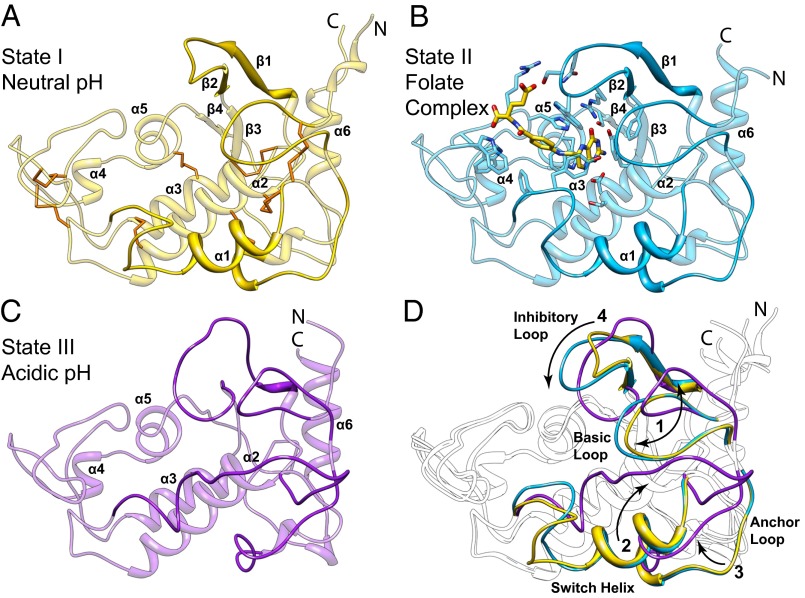
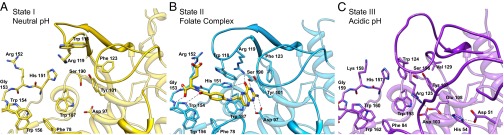
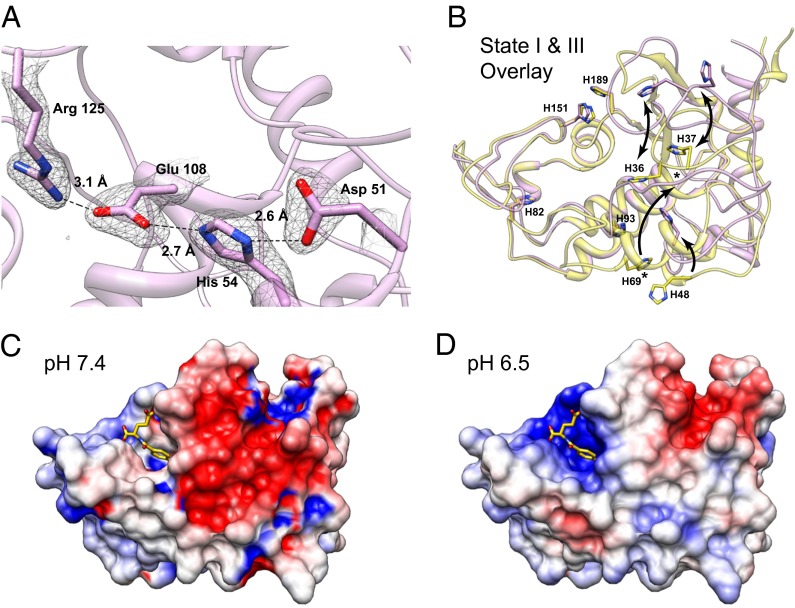
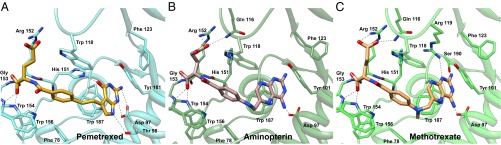
Antifolates, folate analogs that inhibit vitamin B9 (folic acid)-using cellular enzymes, have been used over several decades for the treatment of cancer and inflammatory diseases. Cellular uptake of the antifolates in clinical use occurs primarily via widely expressed facilitative membrane transporters. More recently, human folate receptors (FRs), high affinity receptors that transport folate via endocytosis, have been proposed as targets for the specific delivery of new classes of antifolates or folate conjugates to tumors or sites of inflammation. The development of specific, FR-targeted antifolates would be accelerated if additional biophysical data, particularly structural models of the receptors, were available. Here we describe six distinct crystallographic models that provide insight into biological trafficking of FRs and distinct binding modes of folate and antifolates to these receptors. From comparison of the structures, we delineate discrete structural conformations representative of key stages in the endocytic trafficking of FRs and propose models for pH-dependent conformational changes. Additionally, we describe the molecular details of human FR in complex with three clinically prevalent antifolates, pemetrexed (also Alimta), aminopterin, and methotrexate. On the whole, our data form the basis for rapid design and implementation of unique, FR-targeted, folate-based drugs for the treatment of cancer and inflammatory diseases.
Keywords: isothermal titration calorimetry; targeted drug delivery.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Hoffbrand AV, Weir DG. The history of folic acid. Br J Haematol. 2001;113(3):579–589. - PubMed
-
- Kamen BA, Smith AK. A review of folate receptor alpha cycling and 5-methyltetrahydrofolate accumulation with an emphasis on cell models in vitro. Adv Drug Deliv Rev. 2004;56(8):1085–1097. - PubMed
-
- Kamen BA, Wang MT, Streckfuss AJ, Peryea X, Anderson RG. Delivery of folates to the cytoplasm of MA104 cells is mediated by a surface membrane receptor that recycles. J Biol Chem. 1988;263(27):13602–13609. - PubMed
-
- Matherly LH, Goldman DI. Membrane transport of folates. Vitam Horm. 2003;66:403–456. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
