

Reversal of gene dysregulation in cultured cytotrophoblasts reveals possible causes of preeclampsia
- PMID: 23934129
- PMCID: PMC3999620
- DOI: 10.1172/JCI66966
Reversal of gene dysregulation in cultured cytotrophoblasts reveals possible causes of preeclampsia
Erratum in
- J Clin Invest. 2013 Oct 1;123(10):4541
Abstract
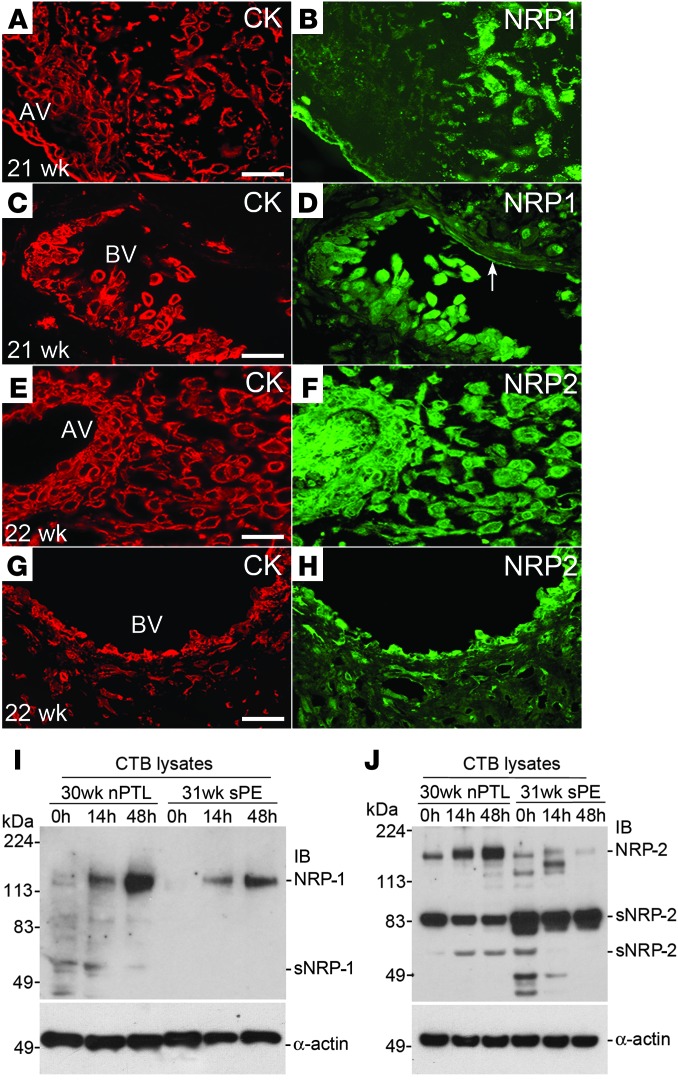
During human pregnancy, a subset of placental cytotrophoblasts (CTBs) differentiates into cells that aggressively invade the uterus and its vasculature, anchoring the progeny and rerouting maternal blood to the placenta. In preeclampsia (PE), CTB invasion is limited, reducing placental perfusion and/or creating intermittent flow. This syndrome, affecting 4%-8% of pregnancies, entails maternal vascular alterations (e.g., high blood pressure, proteinuria, and edema) and, in some patients, fetal growth restriction. The only cure is removal of the faulty placenta, i.e., delivery. Previously, we showed that defective CTB differentiation contributes to the placental component of PE, but the causes were unknown. Here, we cultured CTBs isolated from PE and control placentas for 48 hours, enabling differentiation and invasion. In various severe forms of PE, transcriptomics revealed common aberrations in CTB gene expression immediately after isolation, including upregulation of SEMA3B, which resolved in culture. The addition of SEMA3B to normal CTBs inhibited invasion and recreated aspects of the PE phenotype. Additionally, SEMA3B downregulated VEGF signaling through the PI3K/AKT and GSK3 pathways, effects that were observed in PE CTBs. We propose that, in severe PE, the in vivo environment dysregulates CTB gene expression; the autocrine actions of the upregulated molecules (including SEMA3B) impair CTB differentiation, invasion and signaling; and patient-specific factors determine the signs.
Figures





Comment in
-
Preeclampsia: a link between trophoblast dysregulation and an antiangiogenic state.J Clin Invest. 2013 Jul;123(7):2775-7. doi: 10.1172/JCI70431. Epub 2013 Jun 24. J Clin Invest. 2013. PMID: 23934119 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
