

Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions
- PMID: 23934893
- PMCID: PMC3781963
- DOI: 10.1534/genetics.113.155382
Precise and heritable genome editing in evolutionarily diverse nematodes using TALENs and CRISPR/Cas9 to engineer insertions and deletions
Abstract
Exploitation of custom-designed nucleases to induce DNA double-strand breaks (DSBs) at genomic locations of choice has transformed our ability to edit genomes, regardless of their complexity. DSBs can trigger either error-prone repair pathways that induce random mutations at the break sites or precise homology-directed repair pathways that generate specific insertions or deletions guided by exogenously supplied DNA. Prior editing strategies using site-specific nucleases to modify the Caenorhabditis elegans genome achieved only the heritable disruption of endogenous loci through random mutagenesis by error-prone repair. Here we report highly effective strategies using TALE nucleases and RNA-guided CRISPR/Cas9 nucleases to induce error-prone repair and homology-directed repair to create heritable, precise insertion, deletion, or substitution of specific DNA sequences at targeted endogenous loci. Our robust strategies are effective across nematode species diverged by 300 million years, including necromenic nematodes (Pristionchus pacificus), male/female species (Caenorhabditis species 9), and hermaphroditic species (C. elegans). Thus, genome-editing tools now exist to transform nonmodel nematode species into genetically tractable model organisms. We demonstrate the utility of our broadly applicable genome-editing strategies by creating reagents generally useful to the nematode community and reagents specifically designed to explore the mechanism and evolution of X chromosome dosage compensation. By developing an efficient pipeline involving germline injection of nuclease mRNAs and single-stranded DNA templates, we engineered precise, heritable nucleotide changes both close to and far from DSBs to gain or lose genetic function, to tag proteins made from endogenous genes, and to excise entire loci through targeted FLP-FRT recombination.
Keywords: CRISPR/Cas9 and TALENs; dosage compensation; genome editing; homology-directed repair; nematode species.
Figures


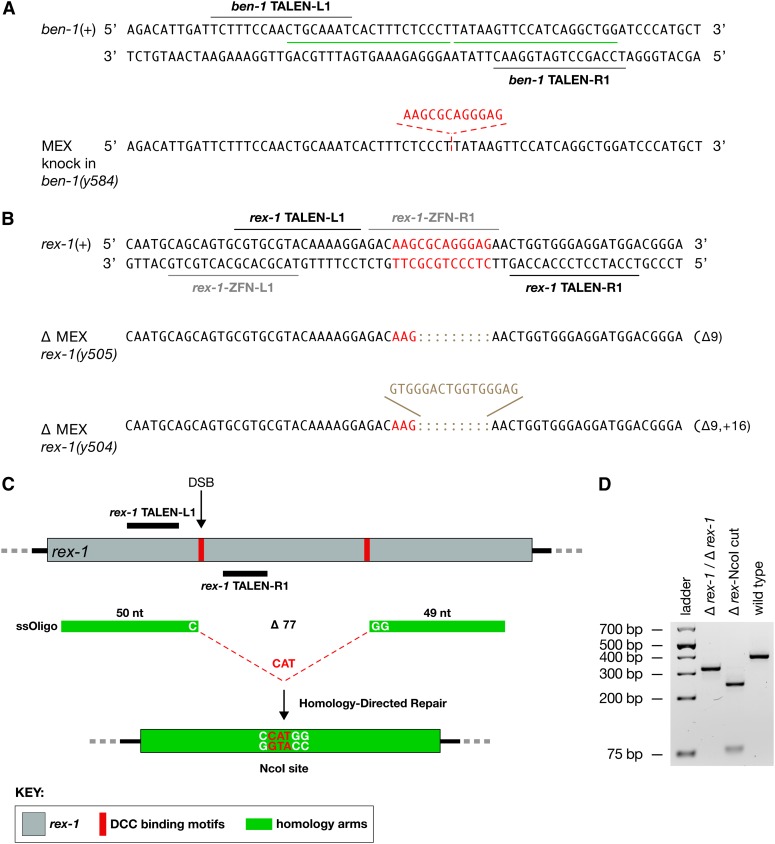
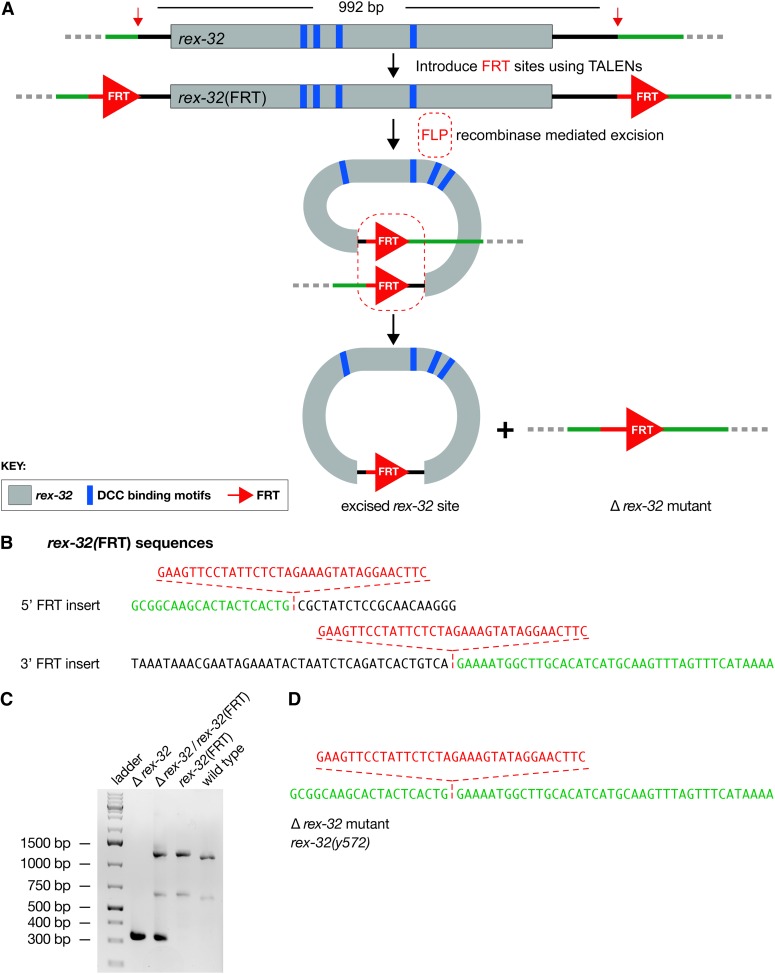
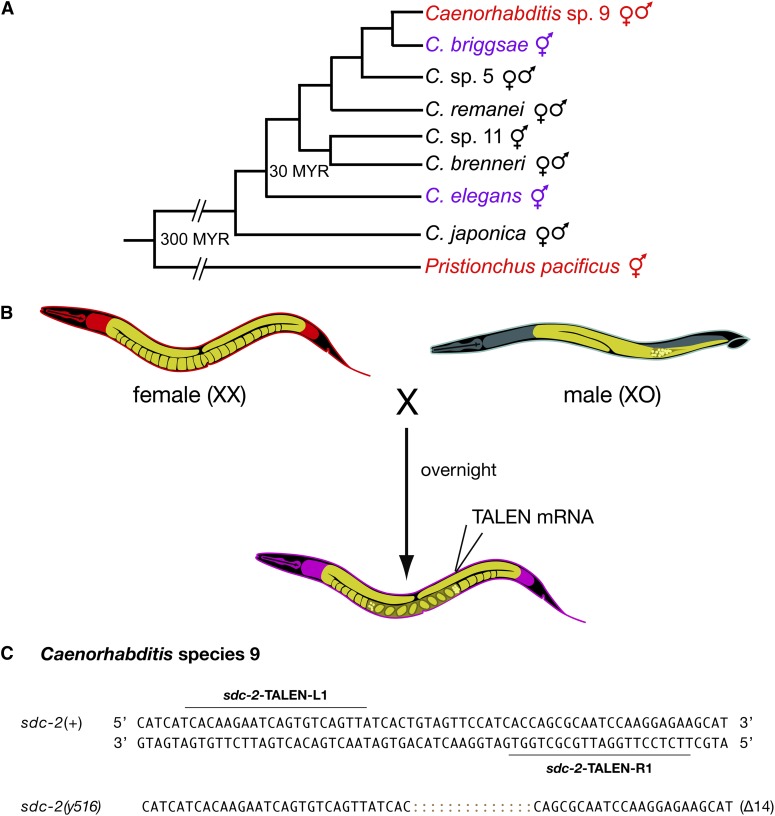
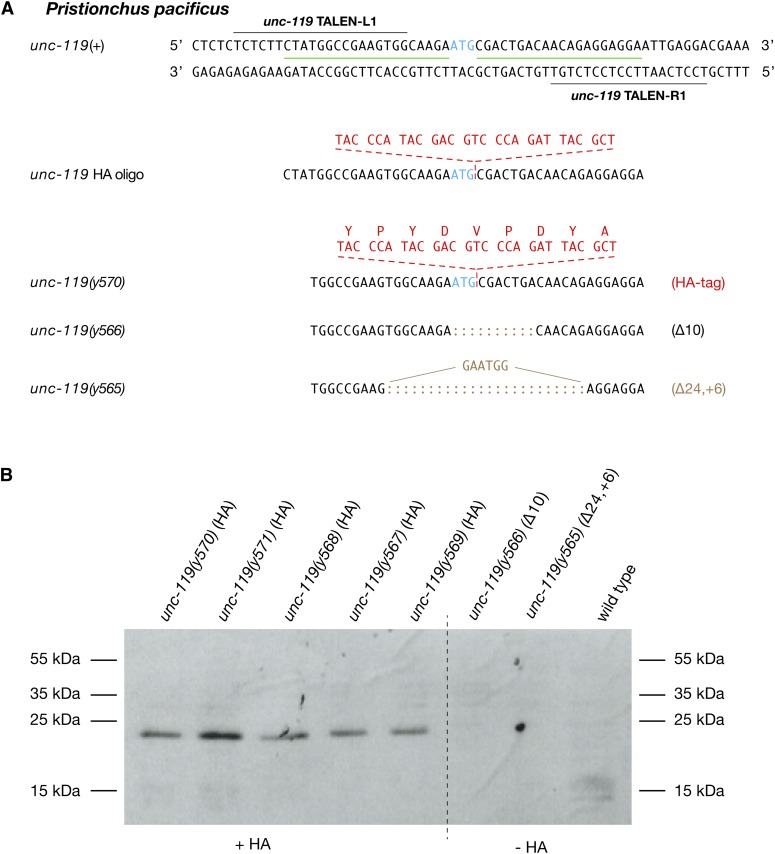
References
-
- Bernard P., Gabant P., Bahassi E. M., Couturier M., 1994. Positive-selection vectors using the F plasmid ccdB killer gene. Gene 148: 71–74. - PubMed
-
- Bhaya D., Davison M., Barrangou R., 2011. CRISPR-Cas systems in bacteria and archaea: versatile small RNAs for adaptive defense and regulation. Annu. Rev. Genet. 45: 273–297. - PubMed
