

A moonlighting enzyme links Escherichia coli cell size with central metabolism
- PMID: 23935518
- PMCID: PMC3723540
- DOI: 10.1371/journal.pgen.1003663
A moonlighting enzyme links Escherichia coli cell size with central metabolism
Abstract
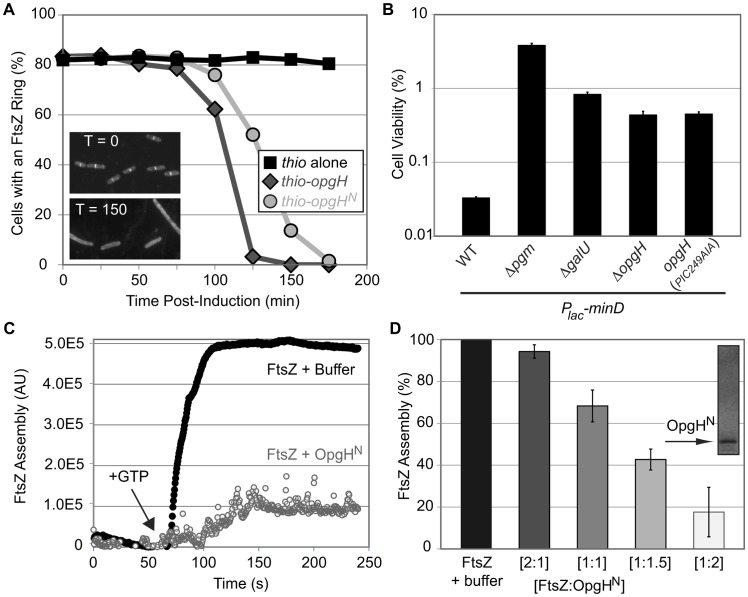
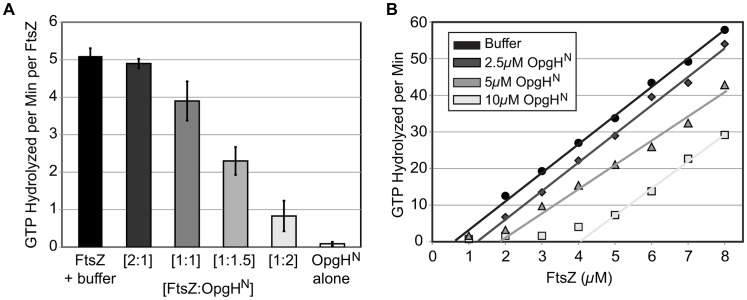
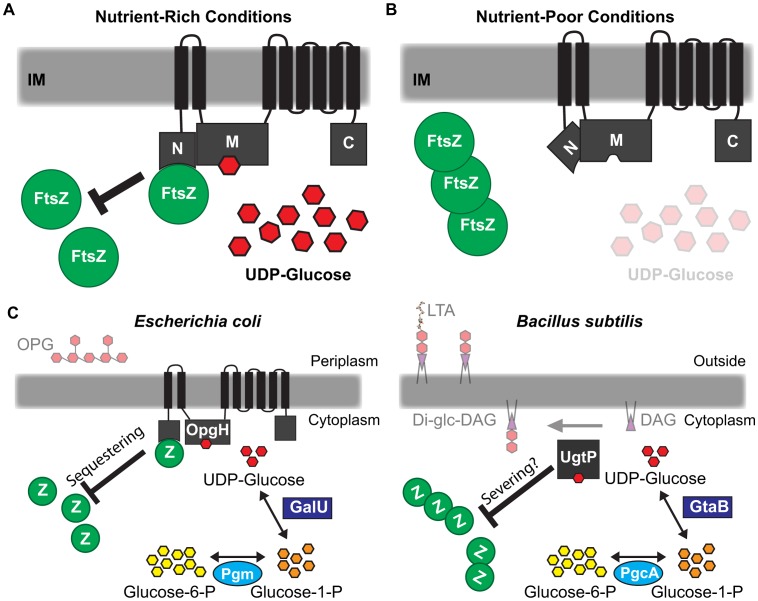
Growth rate and nutrient availability are the primary determinants of size in single-celled organisms: rapidly growing Escherichia coli cells are more than twice as large as their slow growing counterparts. Here we report the identification of the glucosyltransferase OpgH as a nutrient-dependent regulator of E. coli cell size. During growth under nutrient-rich conditions, OpgH localizes to the nascent septal site, where it antagonizes assembly of the tubulin-like cell division protein FtsZ, delaying division and increasing cell size. Biochemical analysis is consistent with OpgH sequestering FtsZ from growing polymers. OpgH is functionally analogous to UgtP, a Bacillus subtilis glucosyltransferase that inhibits cell division in a growth rate-dependent fashion. In a striking example of convergent evolution, OpgH and UgtP share no homology, have distinct enzymatic activities, and appear to inhibit FtsZ assembly through different mechanisms. Comparative analysis of E. coli and B. subtilis reveals conserved aspects of growth rate regulation and cell size control that are likely to be broadly applicable. These include the conservation of uridine diphosphate glucose as a proxy for nutrient status and the use of moonlighting enzymes to couple growth rate-dependent phenomena to central metabolism.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Schaechter M, Maaløe O, Kjeldgaard NO (1958) Dependency on medium and temperature of cell size and chemical composition during balanced grown of Salmonella typhimurium . J Gen Microbiol 19: 592–606. - PubMed
-
- Fantes P, Nurse P (1977) Control of cell size at division in fission yeast by a growth-modulated size control over nuclear division. Exp Cell Res 107: 377–386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
