

The C57BL/6J mouse exhibits sporadic congenital portosystemic shunts
- PMID: 23936100
- PMCID: PMC3720623
- DOI: 10.1371/journal.pone.0069782
The C57BL/6J mouse exhibits sporadic congenital portosystemic shunts
Abstract
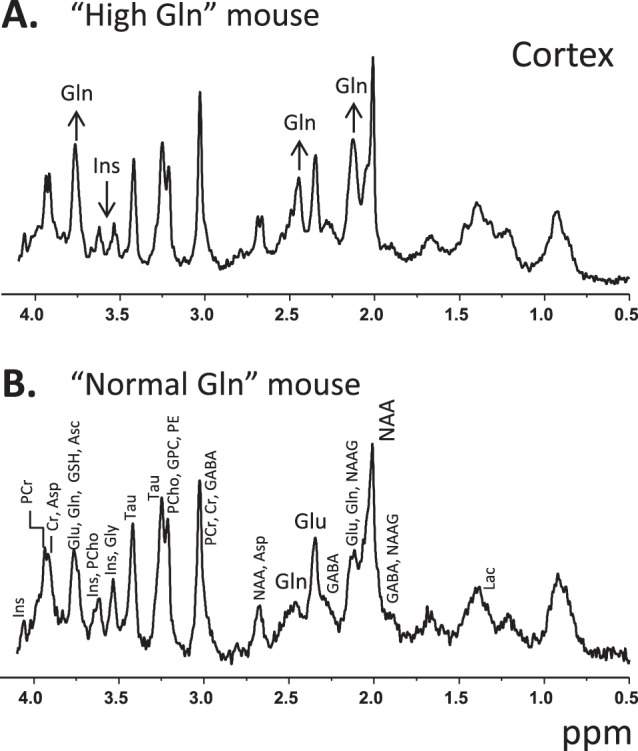
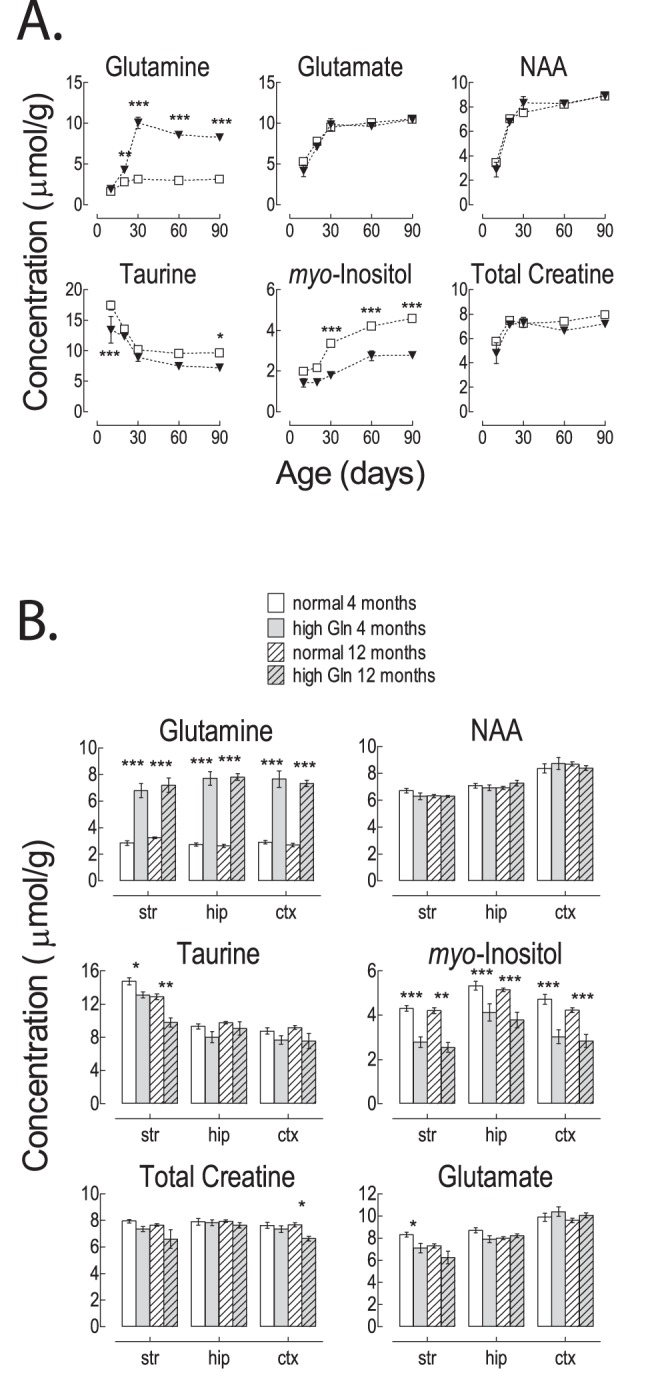
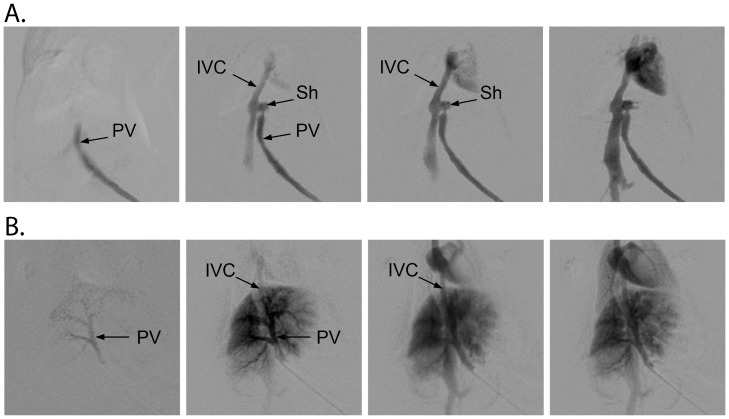
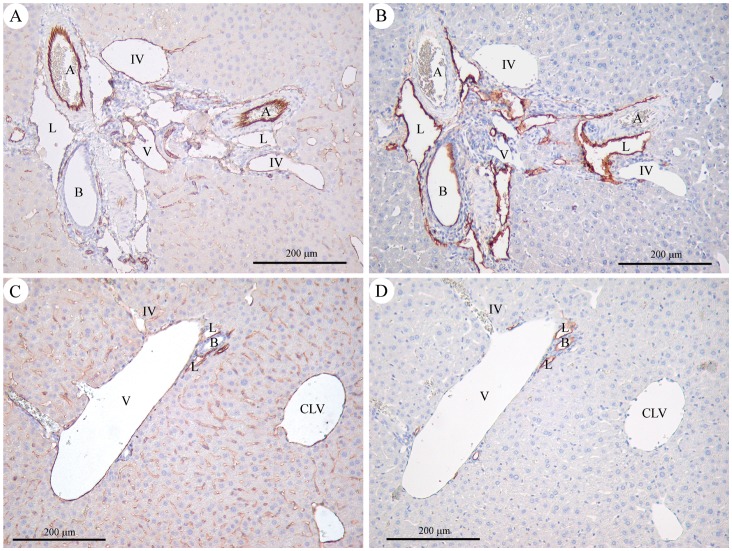
C57BL/6 mice are the most widely used strain of laboratory mice. Using in vivo proton Magnetic Resonance Spectroscopy ((1)H MRS), we have repeatedly observed an abnormal neurochemical profile in the brains of both wild-type and genetically modified mice derived from the C57BL/6J strain, consisting of a several fold increase in cerebral glutamine and two fold decrease in myo-inositol. This strikingly abnormal neurochemical "phenotype" resembles that observed in chronic liver disease or portosystemic shunting and appeared to be independent of transgene, origin or chow and was not associated with liver failure. As many as 25% of animals displayed the abnormal neurochemical profile, questioning the reliability of this model for neurobiology. We conducted an independent study to determine if this neurochemical profile was associated with portosystemic shunting. Our results showed that 100% of the mice with high brain glutamine displayed portosystemic shunting by concomitant portal angiography while all mice with normal brain glutamine did not. Since portosystemic shunting is known to cause alterations in gene expression in many organs including the brain, we conclude that portosystemic shunting may be the most significant problem associated with C57BL/6J inbreeding both for its effect on the central nervous system and for its systemic repercussions.
Conflict of interest statement
Figures
References
-
- Zurita E, Chagoyen M, Cantero M, Alonso R, Gonzalez-Neira A, et al. (2011) Genetic polymorphisms among C57BL/6 mouse inbred strains. Transgenic Res 20: 481–489. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
