

Cell guidance on nanogratings: a computational model of the interplay between PC12 growth cones and nanostructures
- PMID: 23936404
- PMCID: PMC3735603
- DOI: 10.1371/journal.pone.0070304
Cell guidance on nanogratings: a computational model of the interplay between PC12 growth cones and nanostructures
Abstract
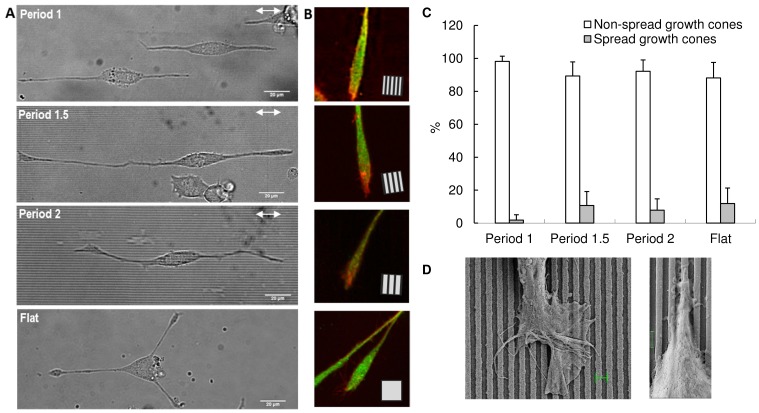
Background: Recently, the effects of nanogratings have been investigated on PC12 with respect to cell polarity, neuronal differentiation, migration, maturation of focal adhesions and alignment of neurites.
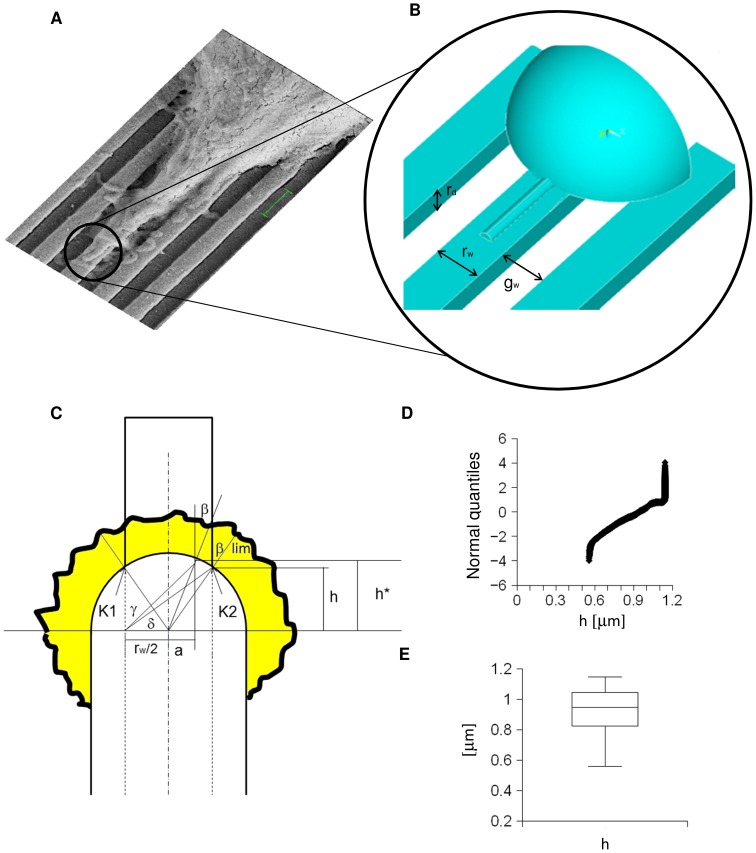
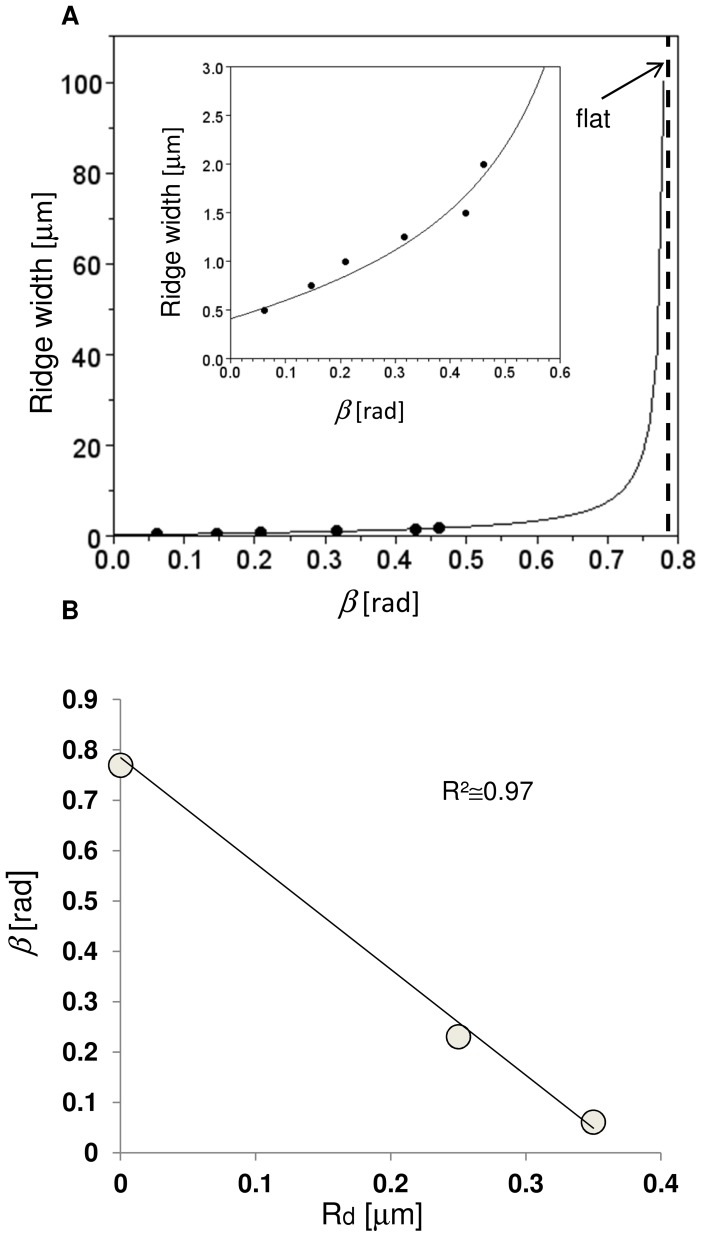
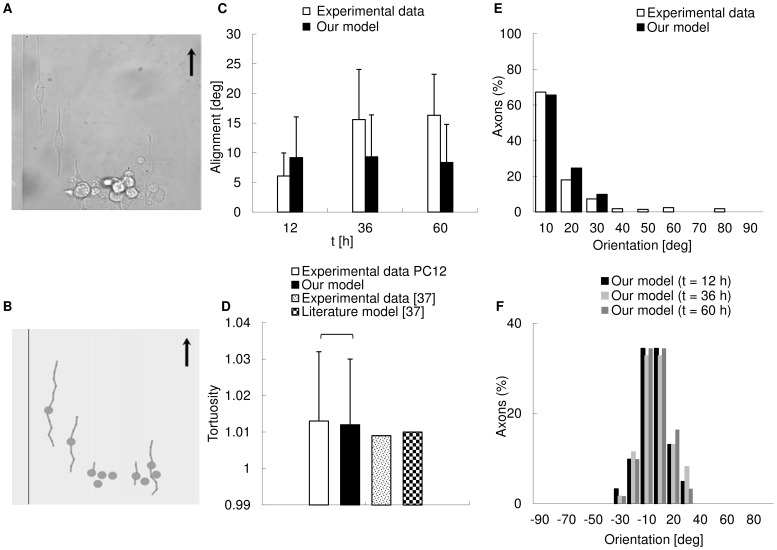
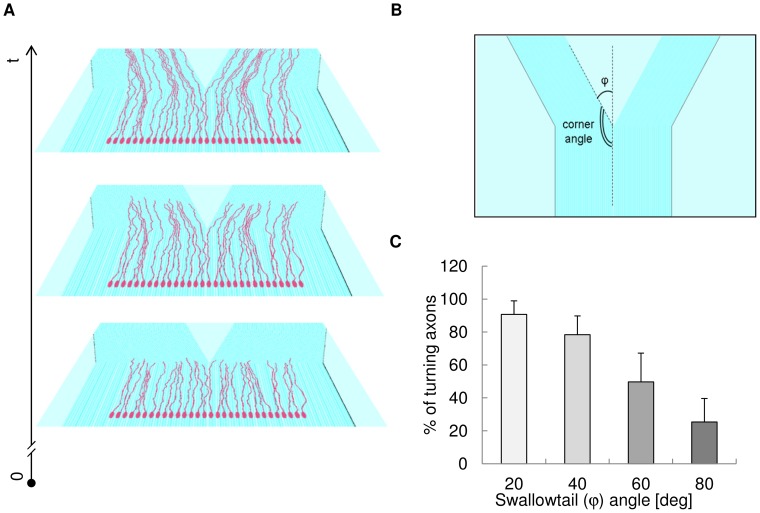
Methodology/principal findings: A synergistic procedure was used to study the mechanism of alignment of PC12 neurites with respect to the main direction of nanogratings. Finite Element simulations were used to qualitatively assess the distribution of stresses at the interface between non-spread growth cones and filopodia, and to study their dependence on filopodial length and orientation. After modelling all adhesions under non-spread growth cone and filopodial protrusions, the values of local stress maxima resulted from the length of filopodia. Since the stress was assumed to be the main triggering cause leading to the increase and stabilization of filopodia, the position of the local maxima was directly related to the orientation of neurites. An analytic closed form equation was then written to quantitatively assess the average ridge width needed to achieve a given neuritic alignment (R(2) = 0.96), and the alignment course, when the ridge depth varied (R(2) = 0.97). A computational framework was implemented within an improved free Java environment (CX3D) and in silico simulations were carried out to reproduce and predict biological experiments. No significant differences were found between biological experiments and in silico simulations (alignment, p = 0.3571; tortuosity, p = 0.2236) with a standard level of confidence (95%).
Conclusions/significance: A mechanism involved in filopodial sensing of nanogratings is proposed and modelled through a synergistic use of FE models, theoretical equations and in silico simulations. This approach shows the importance of the neuritic terminal geometry, and the key role of the distribution of the adhesion constraints for the cell/substrate coupling process. Finally, the effects of the geometry of nanogratings were explicitly considered in cell/surface interactions thanks to the analytic framework presented in this work.
Conflict of interest statement
Figures
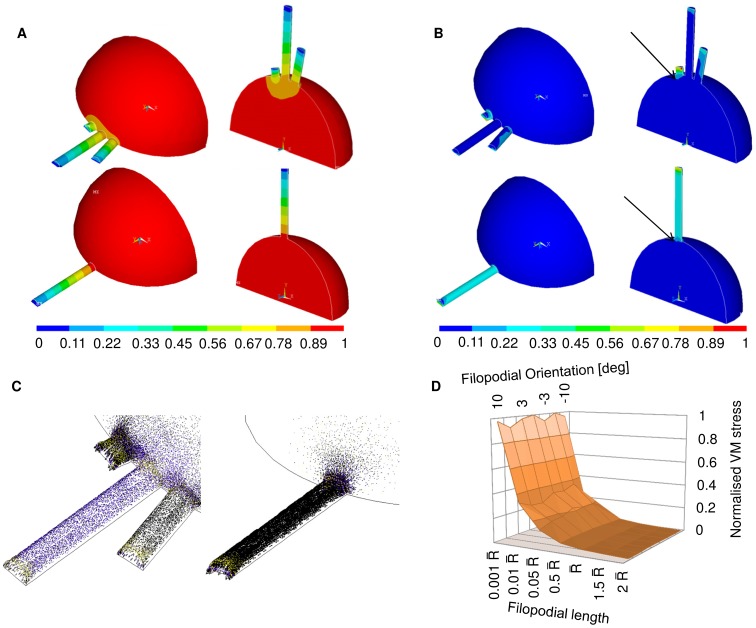
 –2
–2 where
where  was the radius of the non-spread growth cone. All values were normalized on the interface VM stress at 0° for a length of 0.001
was the radius of the non-spread growth cone. All values were normalized on the interface VM stress at 0° for a length of 0.001 .
.
References
-
- Arimura N, Kaibuchi K (2007) Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat Rev Neurosci 8: 194–205. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
