

TACC3 is essential for EGF-mediated EMT in cervical cancer
- PMID: 23936413
- PMCID: PMC3731346
- DOI: 10.1371/journal.pone.0070353
TACC3 is essential for EGF-mediated EMT in cervical cancer
Erratum in
- PLoS One. 2013;8(9). doi:10.1371/annotation/23c9bde1-5ced-4eb9-8a73-c53f3f2913d4. Breuer, Eun-Kyoung Yim [corrected to Breuer, Eun-Kyoung]
Abstract
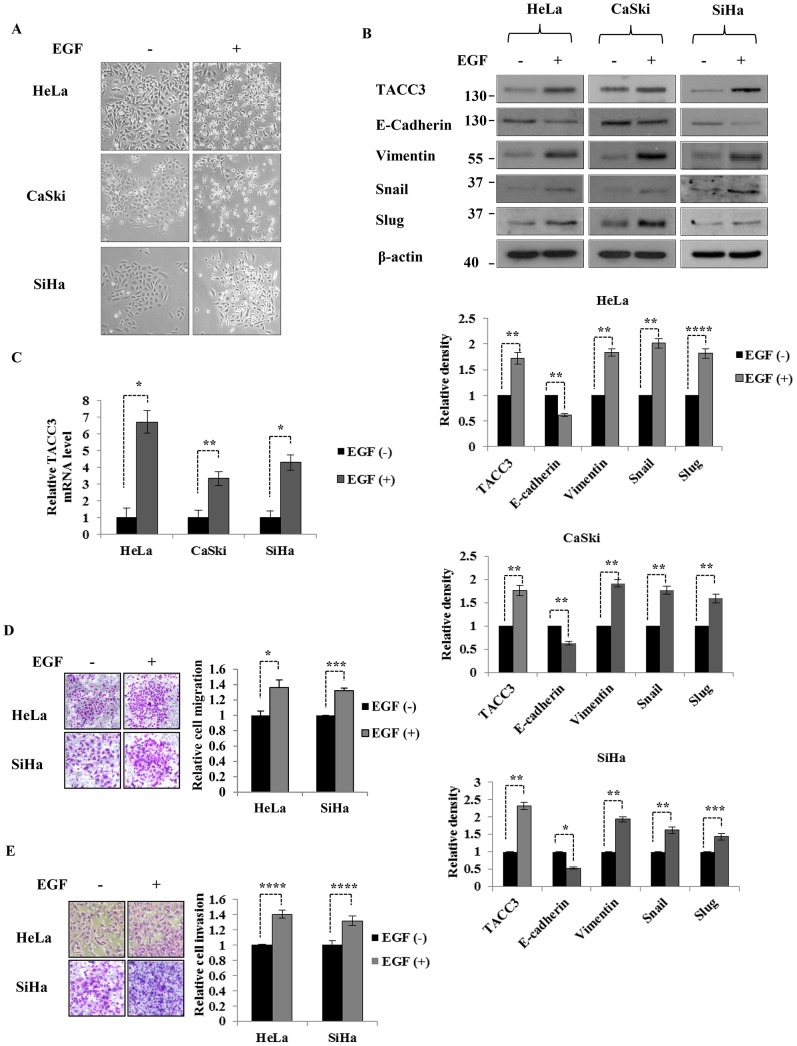
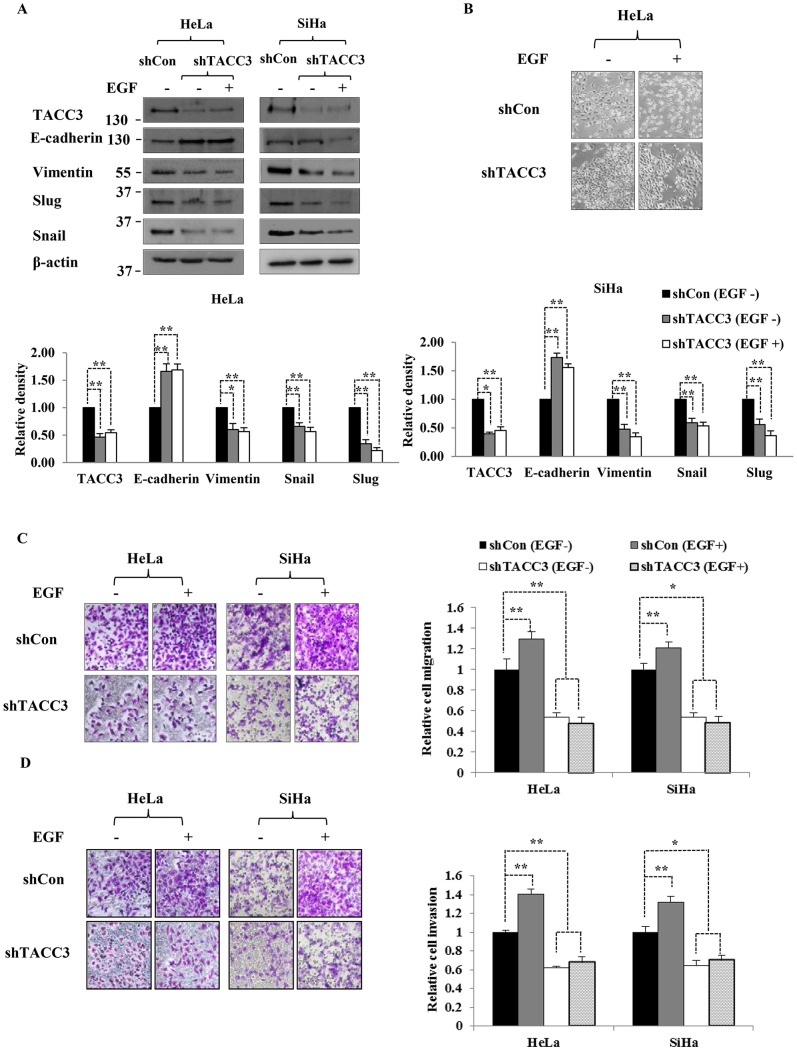
The third member of transforming acidic coiled-coil protein (TACC) family, TACC3, has been shown to be an important player in the regulation of centrosome/microtubule dynamics during mitosis and found to be deregulated in a variety of human malignancies. Our previous studies have suggested that TACC3 may be involved in cervical cancer progression and chemoresistance, and its overexpression can induce epithelial-mesenchymal transition (EMT) by activating the phosphatidylinositol 3-kinase (PI3K)/Akt and extracellular signal-regulated protein kinases (ERKs) signal transduction pathways. However, the upstream mechanisms of TACC3-mediated EMT and its functional/clinical importance in human cervical cancer remain elusive. Epidermal growth factor (EGF) has been shown to be a potent inducer of EMT in cervical cancer and associated with tumor invasion and metastasis. In this study, we found that TACC3 is overexpressed in cervical cancer and can be induced upon EGF stimulation. The induction of TACC3 by EGF is dependent on the tyrosine kinase activity of the EGF receptor (EGFR). Intriguingly, depletion of TACC3 abolishes EGF-mediated EMT, suggesting that TACC3 is required for EGF/EGFR-driven EMT process. Moreover, Snail, a key player in EGF-mediated EMT, is found to be correlated with the expression of TACC3 in cervical cancer. Collectively, our study highlights a novel function for TACC3 in EGF-mediated EMT process and suggests that targeting of TACC3 may be an attractive strategy to treat cervical cancers driven by EGF/EGFR signaling pathways.
Conflict of interest statement
Figures




References
-
- Huber MA, Kraut N, Beug H (2005) Molecular requirements for epithelial-mesenchymal transition during tumor progression. Curr Opin Cell Biol 17: 548–558. - PubMed
-
- Thiery JP, Acloque H, Huang RY, Nieto MA (2009) Epithelial-mesenchymal transitions in development and disease. Cell 139: 871–890. - PubMed
-
- Tanos B, Rodriguez-Boulan E (2008) The epithelial polarity program: machineries involved and their hijacking by cancer. Oncogene 27: 6939–6957. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
