

Crucial role of perilipin-3 (TIP47) in formation of lipid droplets and PGE2 production in HL-60-derived neutrophils
- PMID: 23936516
- PMCID: PMC3731282
- DOI: 10.1371/journal.pone.0071542
Crucial role of perilipin-3 (TIP47) in formation of lipid droplets and PGE2 production in HL-60-derived neutrophils
Abstract
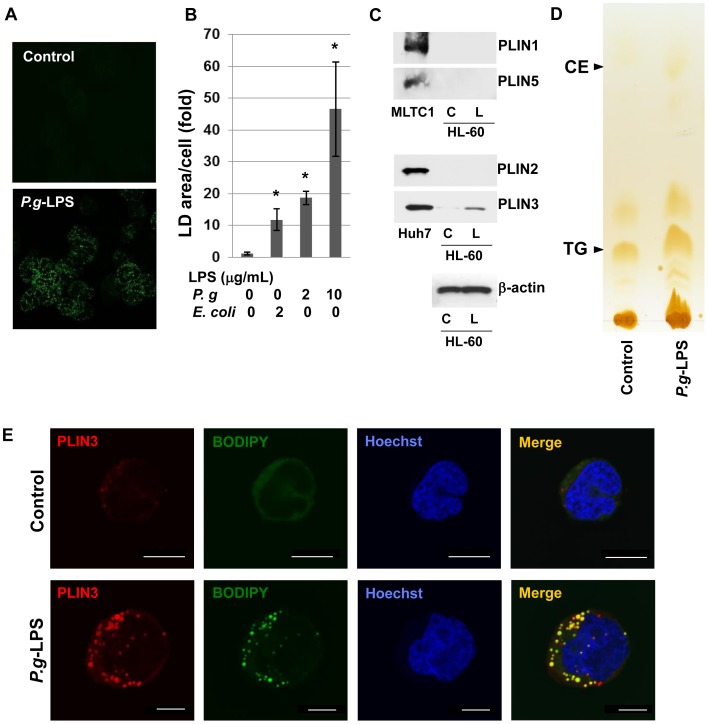
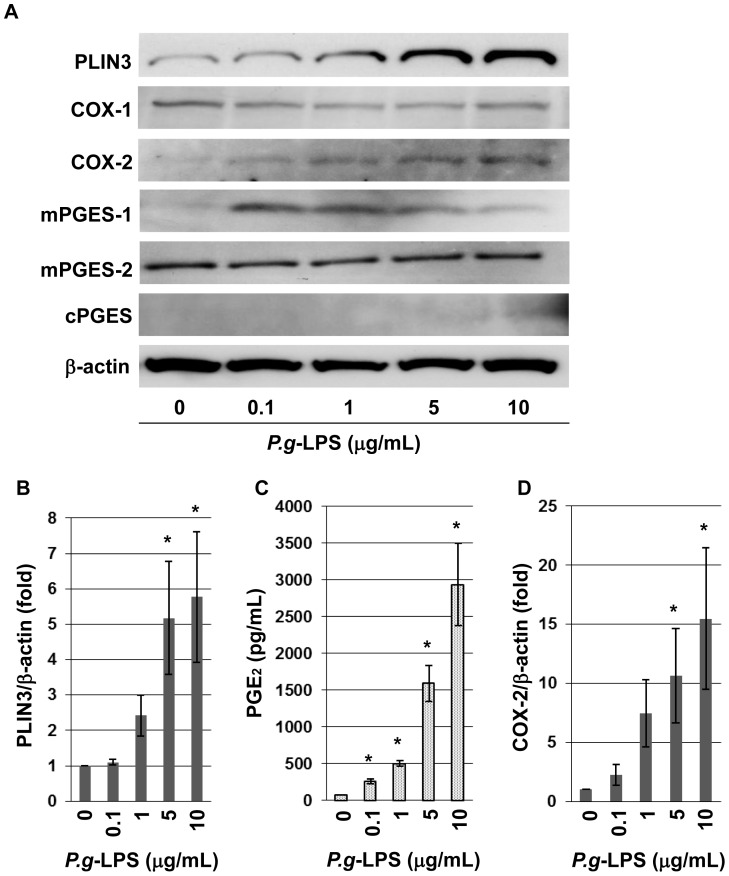
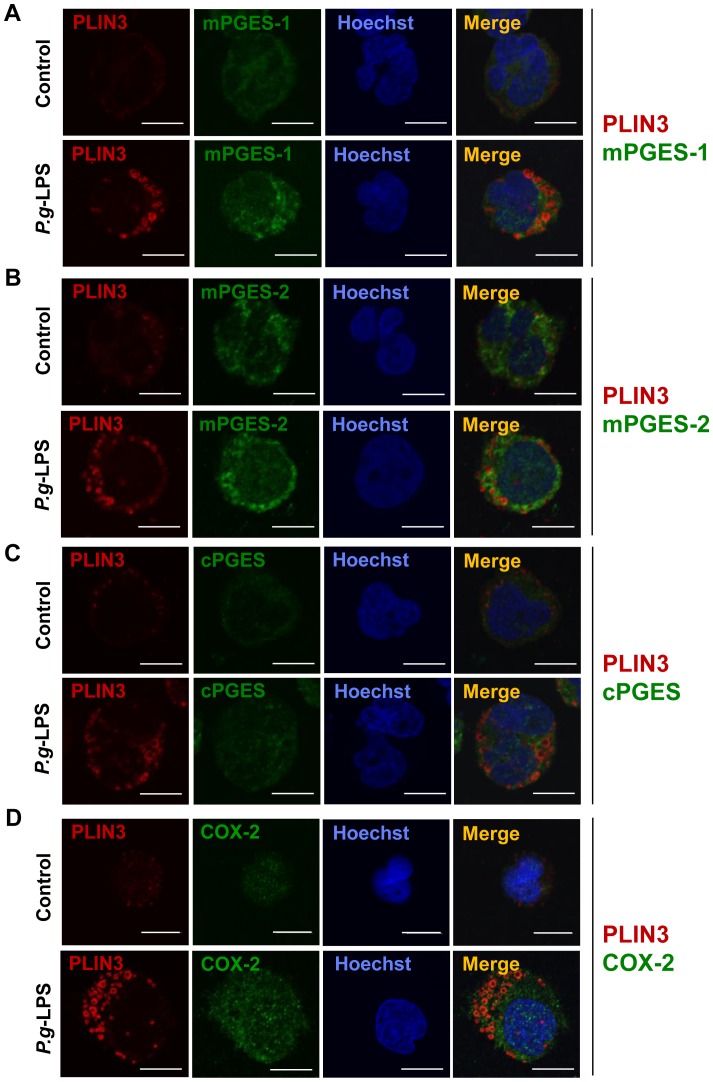
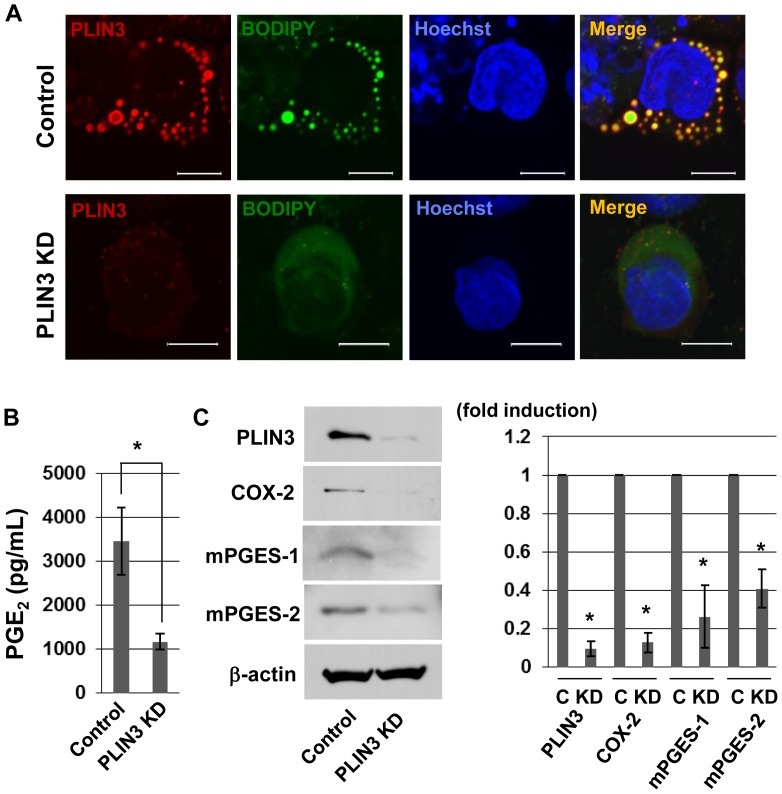
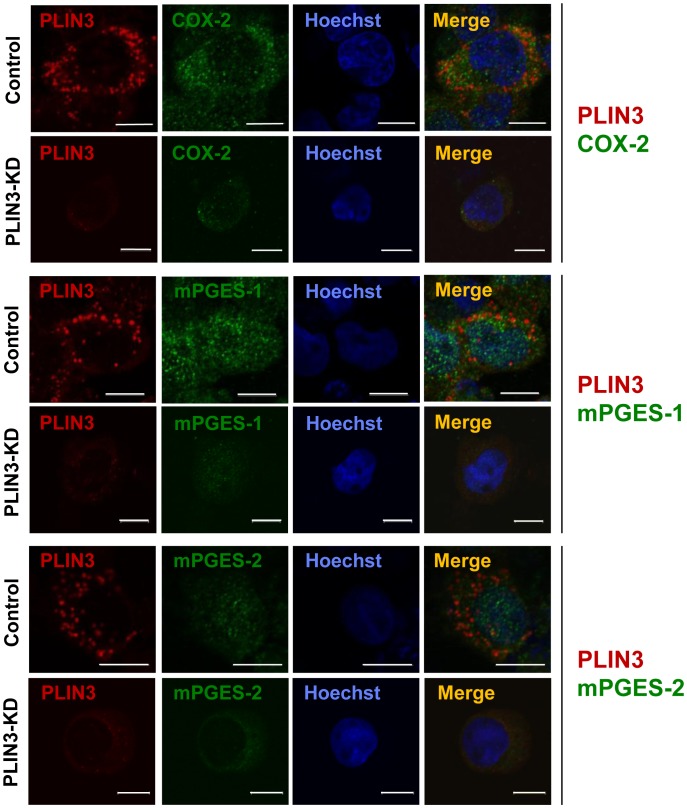
Cytosolic lipid droplets (LDs), which are now recognized as multifunctional organelles, accumulate in leukocytes under various inflammatory conditions. However, little is known about the characteristic features of LDs in neutrophils. In this study, we show that perilipin-3 (PLIN3; formerly called TIP47) is involved in LD formation and the inflammatory response in HL-60-derived neutrophils. HL-60, a promyelocytic cell line, was differentiated into neutrophils via treatment with all-trans retinoic acid. After differentiation, cells were stimulated with Porphyromonas gingivalis lipopolysaccharide (P.g-LPS), a major pathogen in adult periodontitis. When HL-60-derived neutrophils were stimulated with P.g-LPS, LDs increased in both number and size. In the differentiated cells, PLIN3 was induced while PLIN1, PLIN2 and PLIN5 were not detected. PGE2 production and the PLIN3 protein level were increased by the P.g-LPS treatment of the cells in a dose-dependent manner. When PLIN3 was down-regulated with siRNA treatment, LDs essentially disappeared and the level of PGE2 secreted in the cell culture medium decreased by 65%. In addition, the suppression of PLIN3 repressed the PGE2 producing enzymes; i.e., microsomal PGE synthase-1, -2 and cyclooxygenase-2. These findings indicate that PLIN3 has a pivotal role in LD-biogenesis in HL-60-derived neutrophils, and that PLIN3 is associated with the synthesis and secretion of PGE2.
Conflict of interest statement
Figures
References
-
- Tauchi-Sato K, Ozeki S, Houjou T, Taguchi R, Fujimoto T (2002) The surface of lipid droplets is a phospholipid monolayer with a unique fatty acid composition. J Biol Chem 227: 44507–44512. - PubMed
-
- Martin S, Patron RG (2006) Lipid droplets: a unified view of a dynamic organelle. Nat Rev Mol Cell Biol 7: 373–378. - PubMed
-
- Lu X, Gruia-Gray J, Copeland NG, Gilbert DJ, Jenkins NA, et al. (2001) The murine perilipin gene: the lipid droplet-associated perilipins derive from tissue-specific, mRNA splice variants and define a gene family of ancient origin. Mamm Genome 12: 741–749. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
