

Availability of public goods shapes the evolution of competing metabolic strategies
- PMID: 23940318
- PMCID: PMC3761572
- DOI: 10.1073/pnas.1308523110
Availability of public goods shapes the evolution of competing metabolic strategies
Abstract
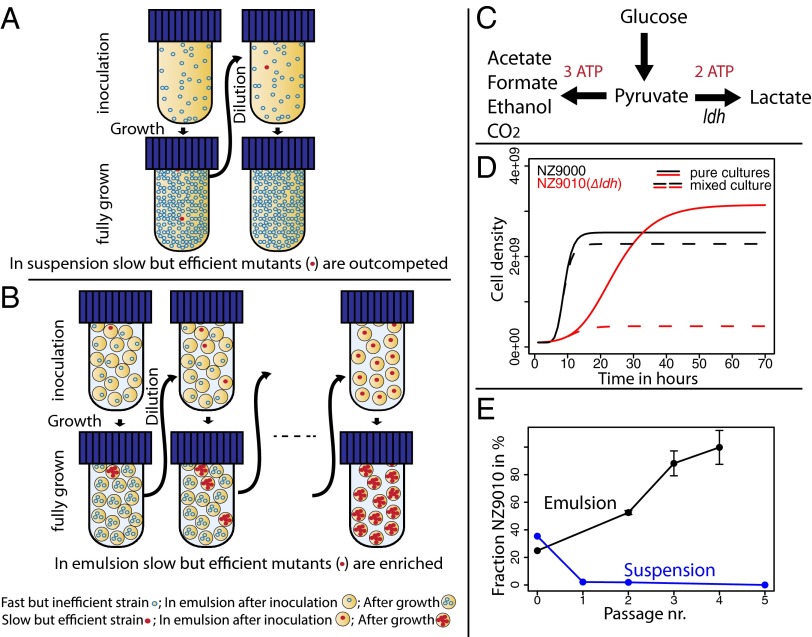
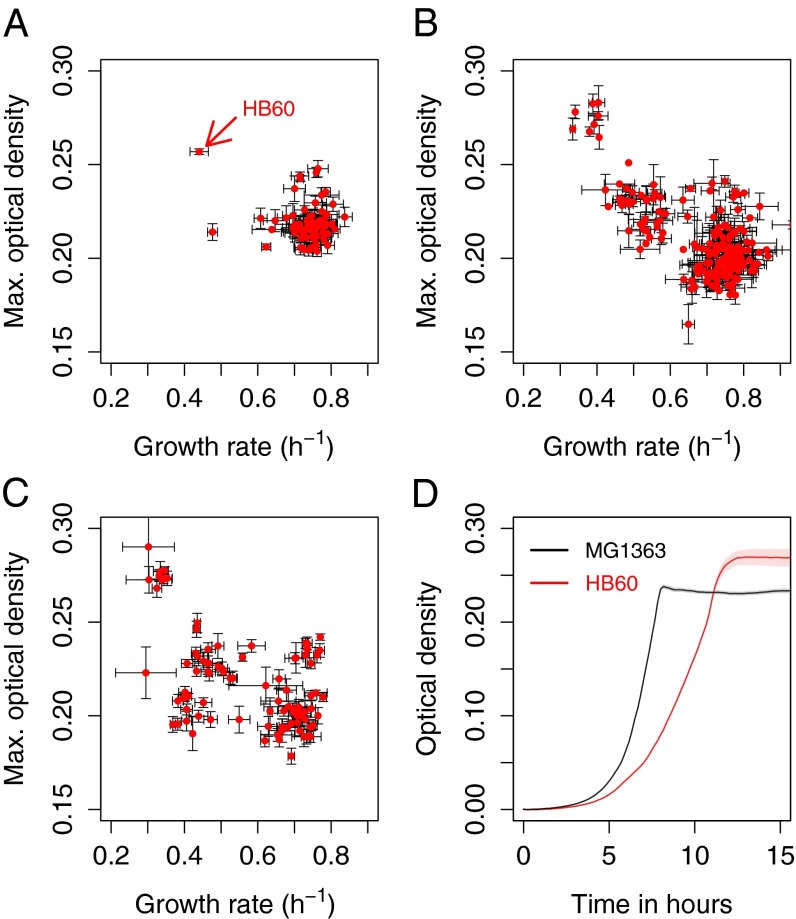
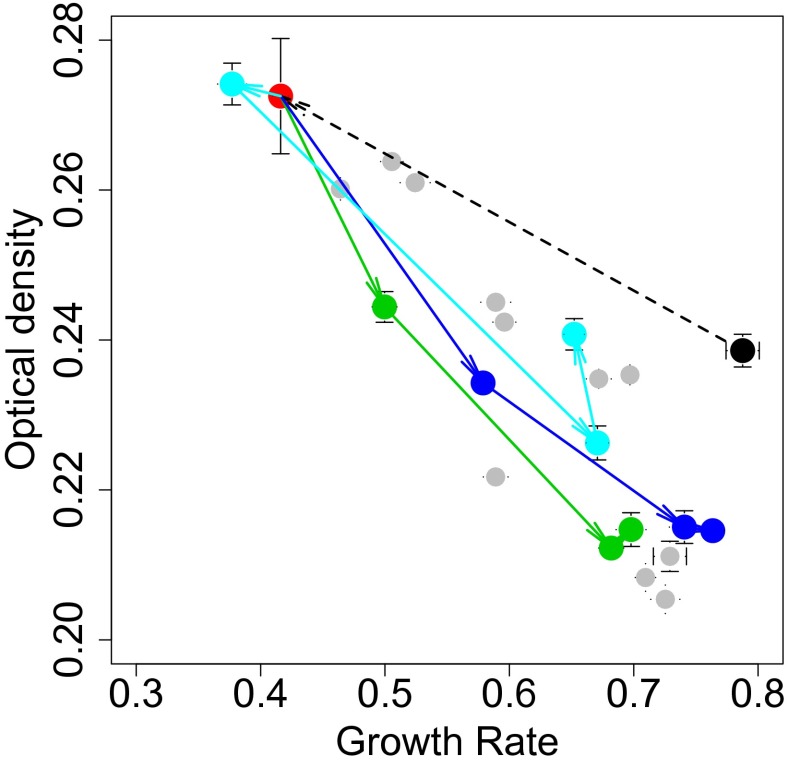
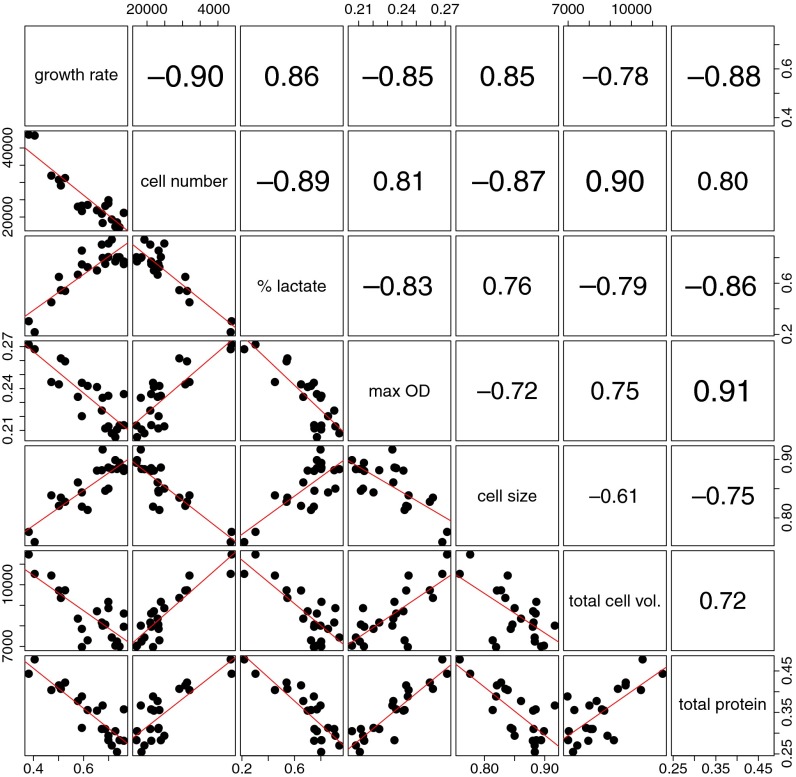
Tradeoffs provide a rationale for the outcome of natural selection. A prominent example is the negative correlation between the growth rate and the biomass yield in unicellular organisms. This tradeoff leads to a dilemma, where the optimization of growth rate is advantageous for an individual, whereas the optimization of the biomass yield would be advantageous for a population. High-rate strategies are observed in a broad variety of organisms such as Escherichia coli, yeast, and cancer cells. Growth in suspension cultures favors fast-growing organisms, whereas spatial structure is of importance for the evolution of high-yield strategies. Despite this realization, experimental methods to directly select for increased yield are lacking. We here show that the serial propagation of a microbial population in a water-in-oil emulsion allows selection of strains with increased biomass yield. The propagation in emulsion creates a spatially structured environment where the growth-limiting substrate is privatized for populations founded by individual cells. Experimental evolution of several isogenic Lactococcus lactis strains demonstrated the existence of a tradeoff between growth rate and biomass yield as an apparent Pareto front. The underlying mutations altered glucose transport and led to major shifts between homofermentative and heterofermentative metabolism, accounting for the changes in metabolic efficiency. The results demonstrated the impact of privatizing a public good on the evolutionary outcome between competing metabolic strategies. The presented approach allows the investigation of fundamental questions in biology such as the evolution of cooperation, cell-cell interactions, and the relationships between environmental and metabolic constraints.
Keywords: droplets; group selection; metabolic engineering; microbial diversity; r/K selection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Beldade P, Koops K, Brakefield PM. Developmental constraints versus flexibility in morphological evolution. Nature. 2002;416(6883):844–847. - PubMed
-
- Velicer GJ, Lenski RE. Evolutionary trade-offs under conditions of resource abundance and scarcity: Experiments with bacteria. Ecology. 1999;80:1168.
-
- Fitzsimmons JM, Schoustra SE, Kerr JT, Kassen R. Population consequences of mutational events: Effects of antibiotic resistance on the r/K trade-off. Evol Ecol. 2009;24:227–236.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
