

Human NAIP and mouse NAIP1 recognize bacterial type III secretion needle protein for inflammasome activation
- PMID: 23940371
- PMCID: PMC3761597
- DOI: 10.1073/pnas.1306376110
Human NAIP and mouse NAIP1 recognize bacterial type III secretion needle protein for inflammasome activation
Abstract
Inflammasome mediated by central nucleotide-binding and oligomerization domain (NOD)-like receptor (NLR) protein is critical for defense against bacterial infection. Here we show that type III secretion system (T3SS) needle proteins from several bacterial pathogens, including Salmonella typhimurium, enterohemorrhagic Escherichia coli, Shigella flexneri, and Burkholderia spp., can induce robust inflammasome activation in both human monocyte-derived and mouse bone marrow macrophages. Needle protein activation of human NRL family CARD domain containing 4 (NLRC4) inflammasome requires the sole human neuronal apoptosis inhibitory protein (hNAIP). Among the seven mouse NAIPs, NAIP1 functions as the mouse counterpart of hNAIP. We found that NAIP1 recognition of T3SS needle proteins was more robust in mouse dendritic cells than in bone marrow macrophages. Needle proteins, as well as flagellin and rod proteins from five different bacteria, exhibited differential and cell type-dependent inflammasome-stimulating activity. Comprehensive profiling of the three types of NAIP ligands revealed that NAIP1 sensing of the needle protein dominated S. flexneri-induced inflammasome activation, particularly in dendritic cells. hNAIP/NAIP1 and NAIP2/5 formed a large oligomeric complex with NLRC4 in the presence of corresponding bacterial ligands, and could support reconstitution of the NLRC4 inflammasome in a ligand-specific manner.
Keywords: NOD-like protein; caspase-1; innate immunity; pathogen-associated molecular pattern; pyroptosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
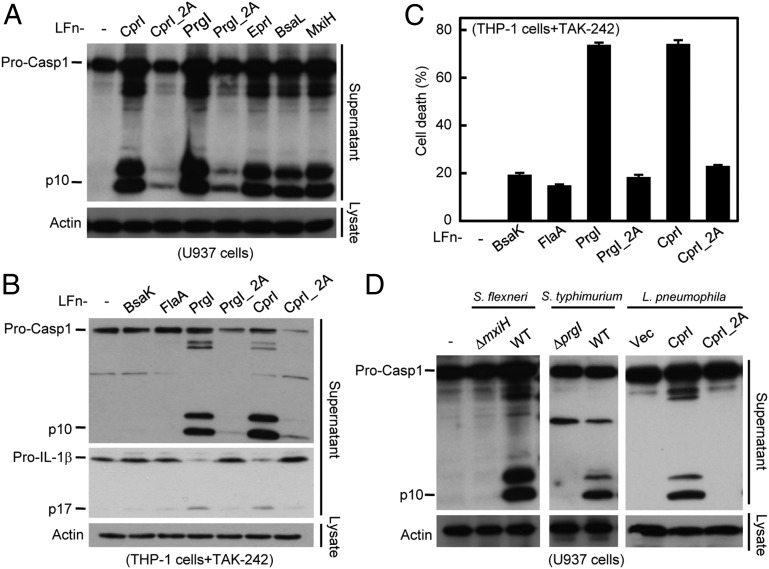
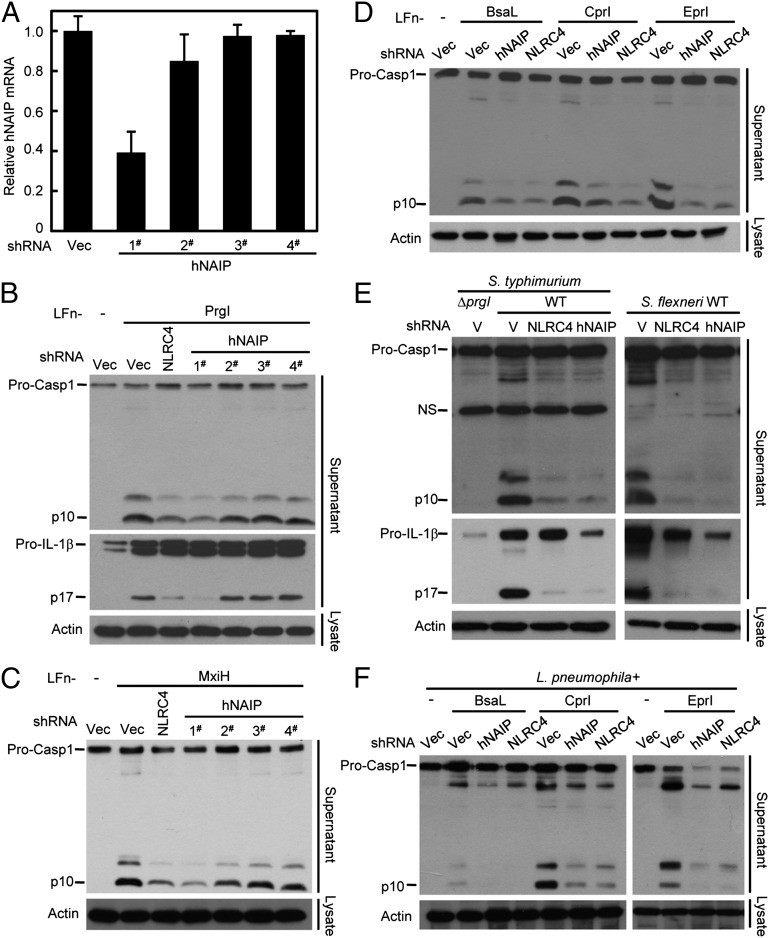
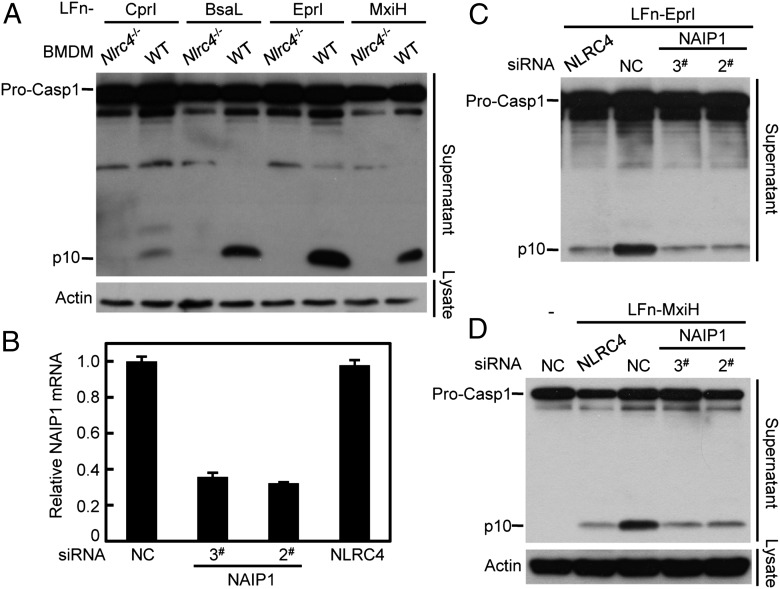
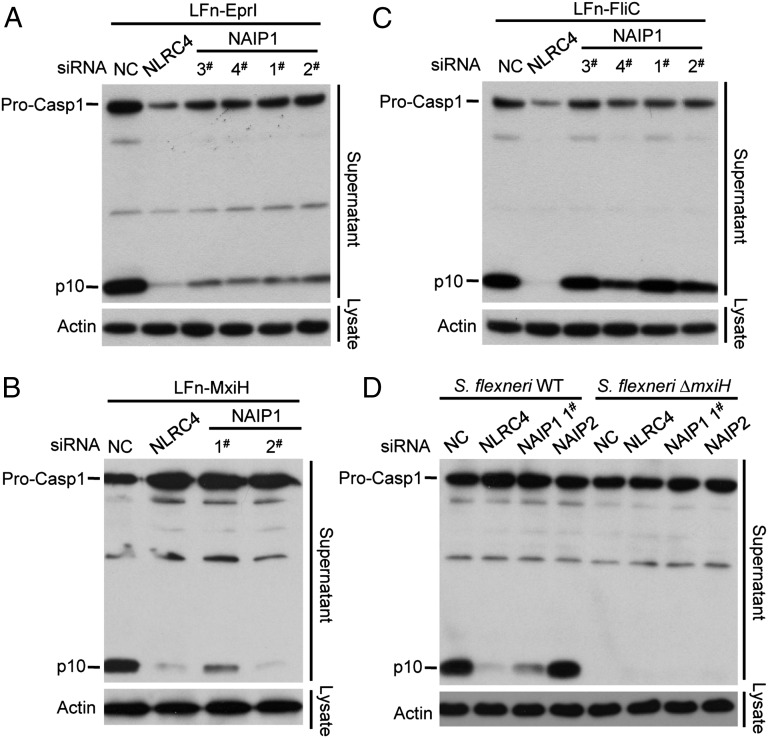

References
-
- Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–820. - PubMed
-
- Chen G, Shaw MH, Kim YG, Nuñez G. NOD-like receptors: Role in innate immunity and inflammatory disease. Annu Rev Pathol. 2009;4:365–398. - PubMed
-
- Ting JP, Kastner DL, Hoffman HM. CATERPILLERs, pyrin and hereditary immunological disorders. Nat Rev Immunol. 2006;6(3):183–195. - PubMed
-
- Schroder K, Tschopp J. The inflammasomes. Cell. 2010;140(6):821–832. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
