

let-7 and miR-140 microRNAs coordinately regulate skeletal development
- PMID: 23940373
- PMCID: PMC3761644
- DOI: 10.1073/pnas.1302797110
let-7 and miR-140 microRNAs coordinately regulate skeletal development
Abstract
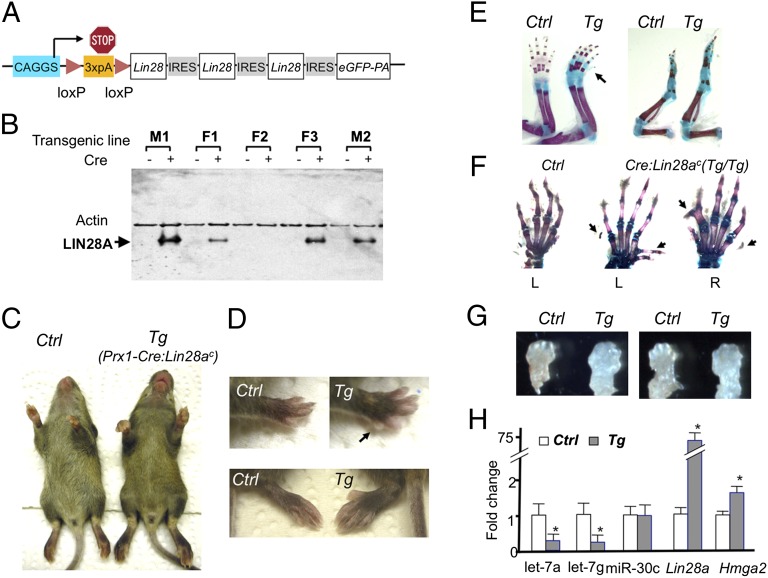
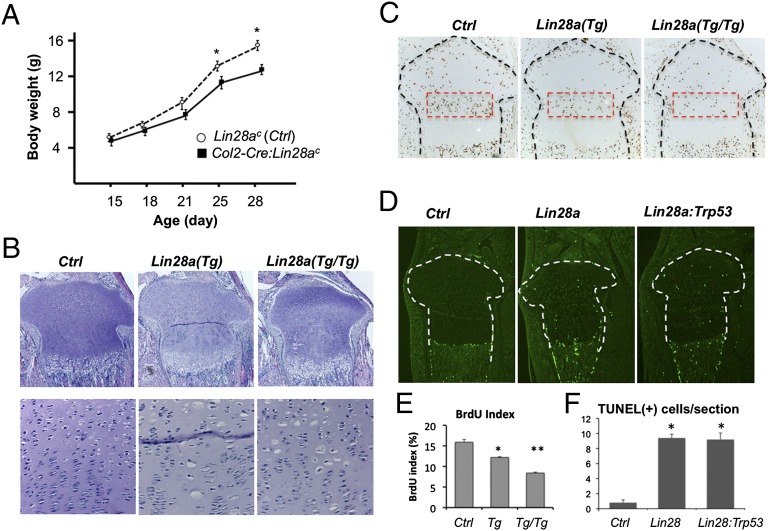
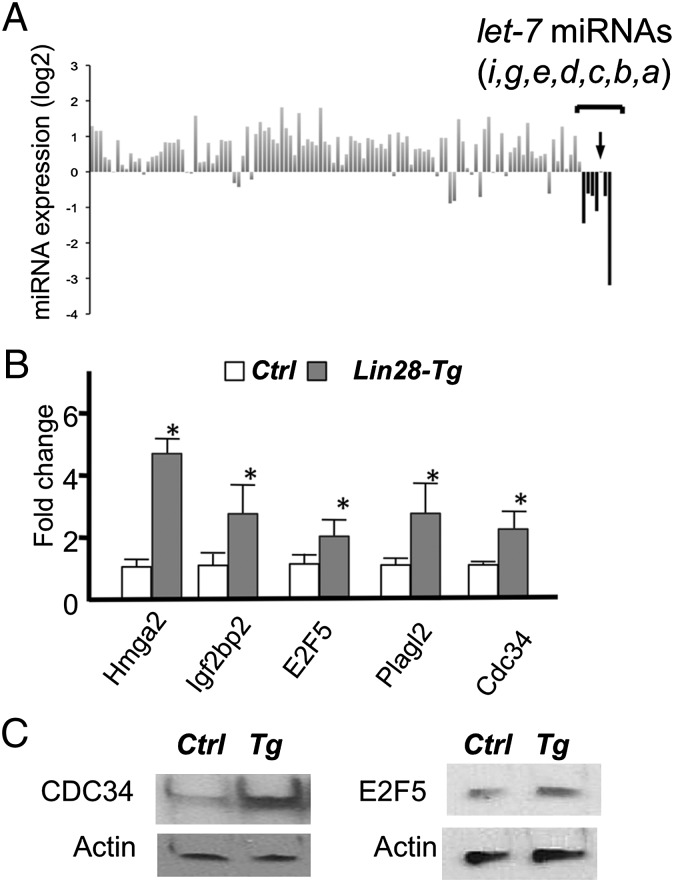
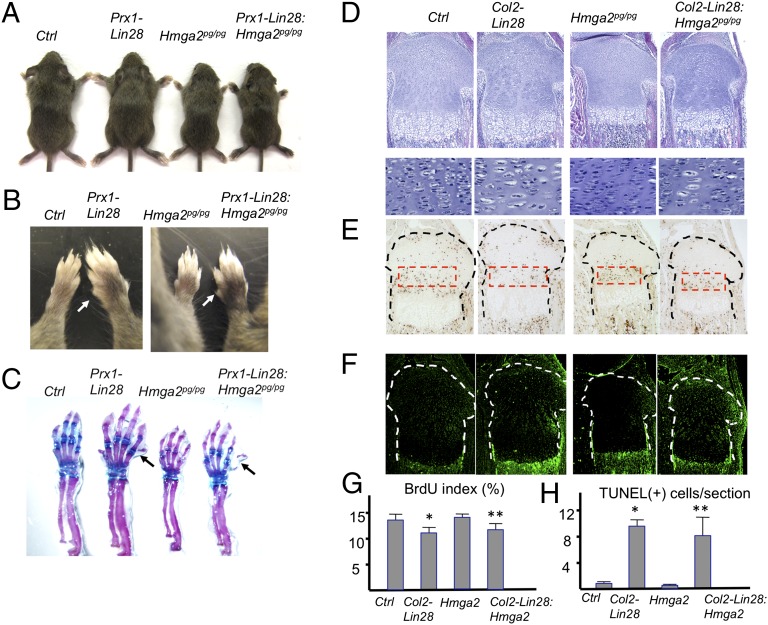
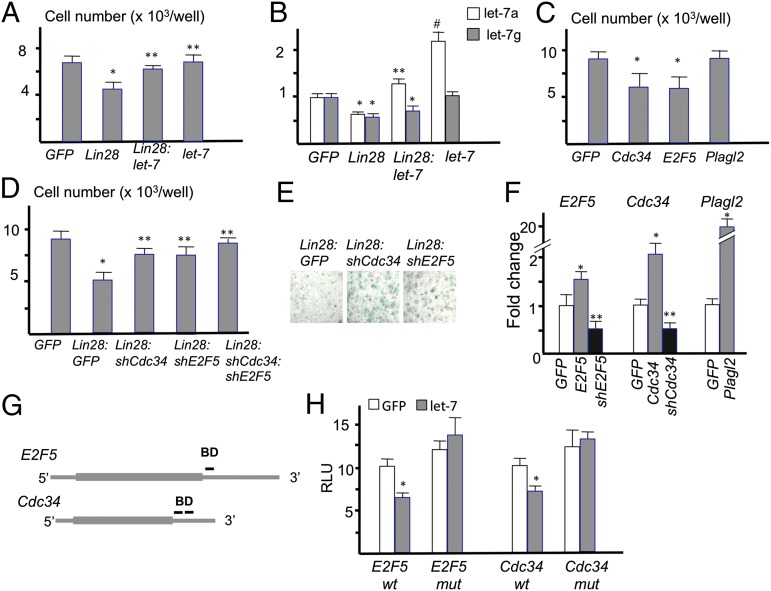
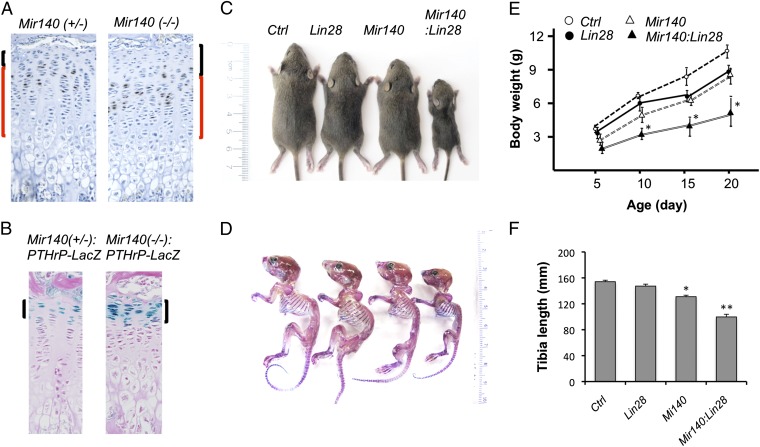
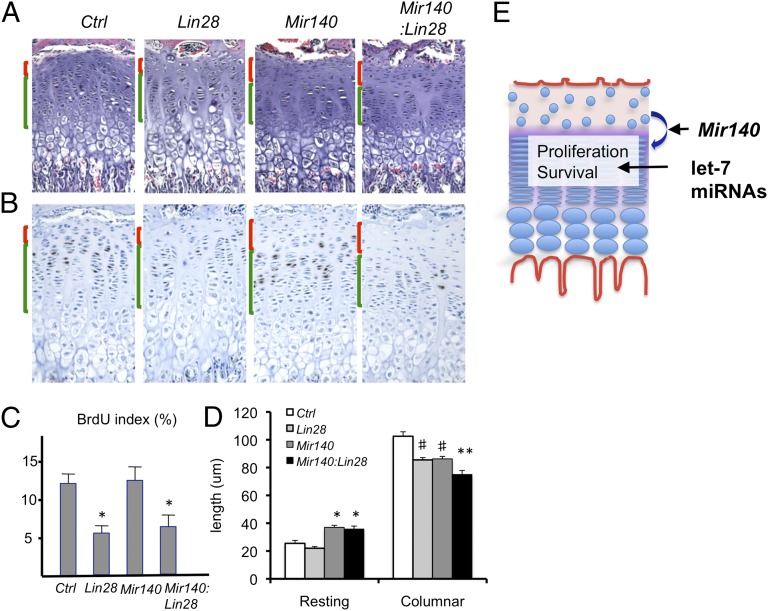
MicroRNAs (miRNAs) play critical roles in multiple processes of skeletal development. A global reduction of miRNAs in growth plate chondrocytes results in defects in both proliferation and differentiation; however, specific microRNAs responsible for these defects have not been identified. In this study, we provide evidence that let-7 miRNAs and microRNA-140 (miR-140), among other miRNAs expressed in chondrocytes, play major roles in endochondral bone development. We overexpressed lin-28 homolog A (Lin28a) to inhibit let-7 miRNA biogenesis in growth plate chondrocytes. Lin28a overexpression efficiently and specifically reduced let-7 miRNAs and up-regulated let-7 target genes. However, unlike the previous notion that let-7 miRNAs inhibit proliferation and growth, suppression of let-7 miRNAs via Lin28a overexpression decreased proliferation in growth plate chondrocytes, likely through up-regulation of the let-7 target cell cycle regulators cell division cycle 34 (Cdc34) and E2F transcription factor 5 (E2F5). Deficiency of the chondrocyte-specific miRNA, miR-140, causes a differentiation defect in growth plate chondrocytes. Although either Lin28a overexpression or miR-140 deficiency alone caused only mild growth impairment, mice with both miR-140 deficiency and Lin28a overexpression in chondrocytes showed a dramatic growth defect. Deregulation of distinct processes in the absence of these miRNAs synergistically decreased the proliferating chondrocyte mass; miR-140 deficiency reduced differentiation into proliferating chondrocytes, whereas Lin28a overexpression decreased proliferation per se.
Keywords: chondrocyte differentiation; chondrocyte proliferation; mouse.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
