

Toll-like receptor-mediated IRE1α activation as a therapeutic target for inflammatory arthritis
- PMID: 23942232
- PMCID: PMC3770952
- DOI: 10.1038/emboj.2013.183
Toll-like receptor-mediated IRE1α activation as a therapeutic target for inflammatory arthritis
Abstract
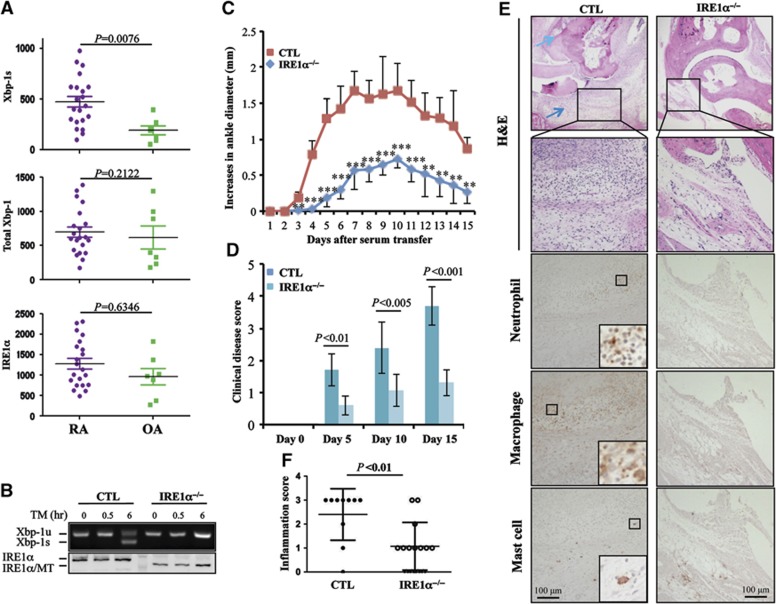
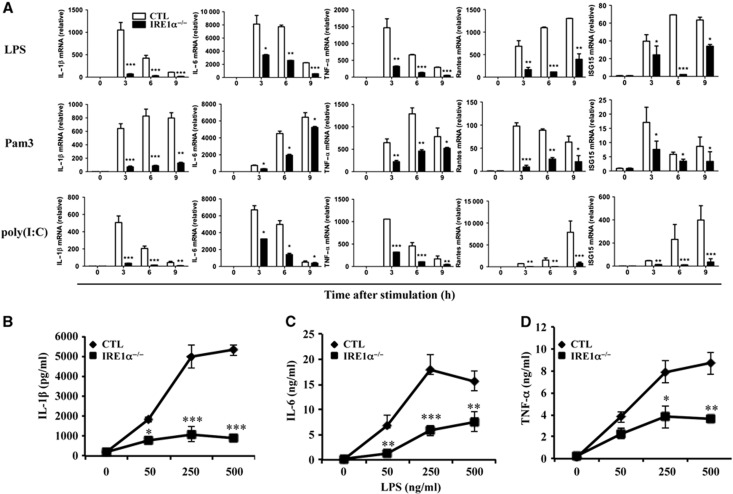
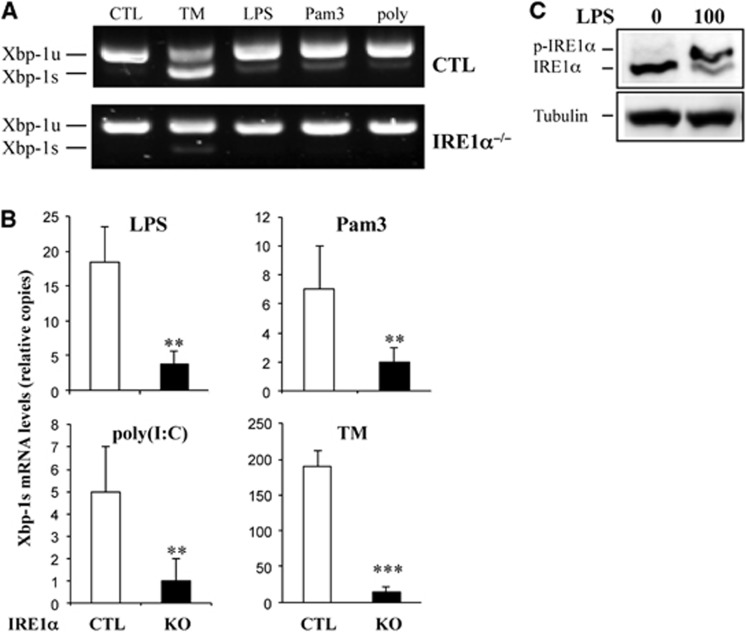
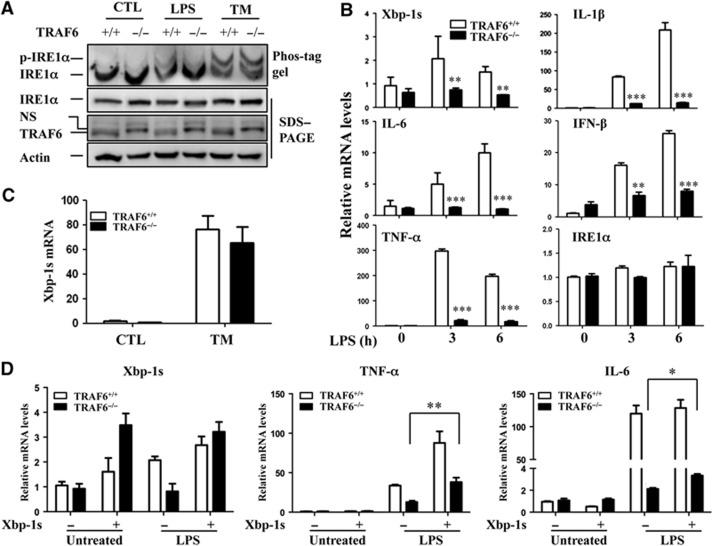
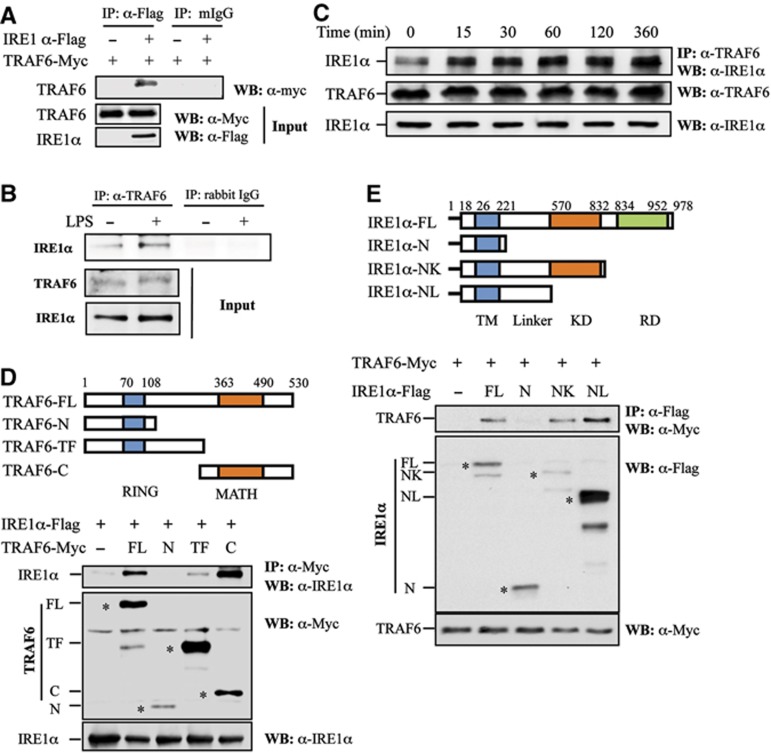
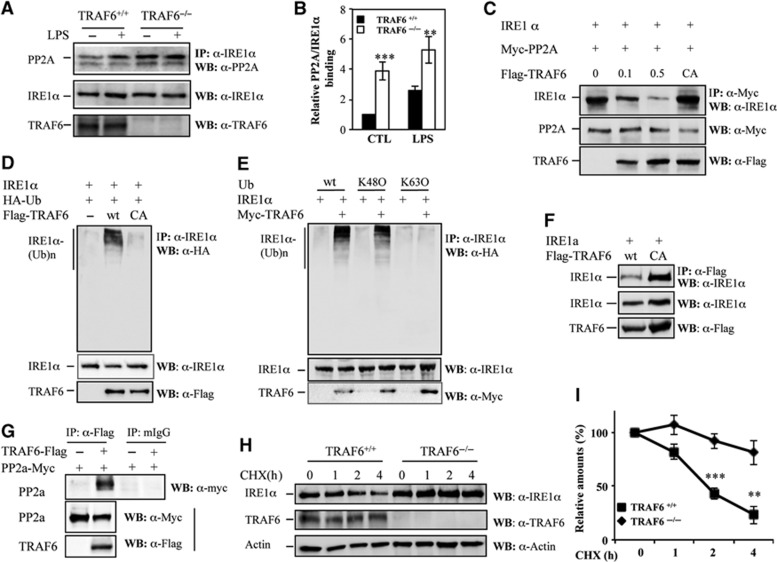
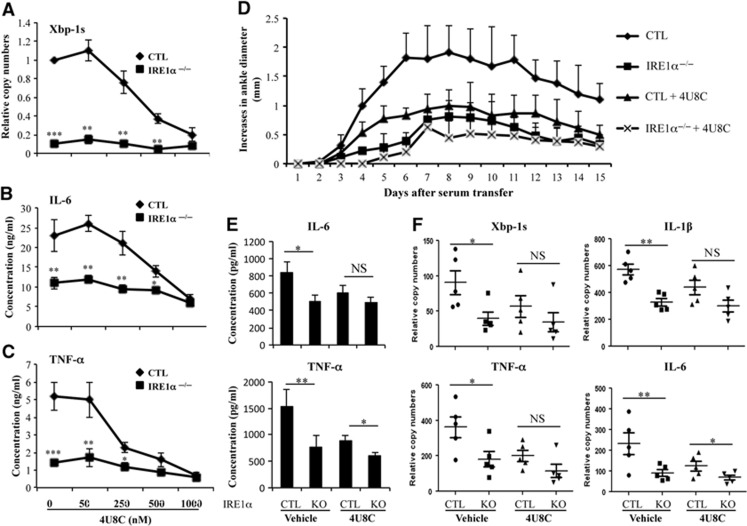
In rheumatoid arthritis (RA), macrophage is one of the major sources of inflammatory mediators. Macrophages produce inflammatory cytokines through toll-like receptor (TLR)-mediated signalling during RA. Herein, we studied macrophages from the synovial fluid of RA patients and observed a significant increase in activation of inositol-requiring enzyme 1α (IRE1α), a primary unfolded protein response (UPR) transducer. Myeloid-specific deletion of the IRE1α gene protected mice from inflammatory arthritis, and treatment with the IRE1α-specific inhibitor 4U8C attenuated joint inflammation in mice. IRE1α was required for optimal production of pro-inflammatory cytokines as evidenced by impaired TLR-induced cytokine production in IRE1α-null macrophages and neutrophils. Further analyses demonstrated that tumour necrosis factor (TNF) receptor-associated factor 6 (TRAF6) plays a key role in TLR-mediated IRE1α activation by catalysing IRE1α ubiquitination and blocking the recruitment of protein phosphatase 2A (PP2A), a phosphatase that inhibits IRE1α phosphorylation. In summary, we discovered a novel regulatory axis through TRAF6-mediated IRE1α ubiquitination in regulating TLR-induced IRE1α activation in pro-inflammatory cytokine production, and demonstrated that IRE1α is a potential therapeutic target for inflammatory arthritis.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Asea A, Rehli M, Kabingu E, Boch JA, Bare O, Auron PE, Stevenson MA, Calderwood SK (2002) Novel signal transduction pathway utilized by extracellular HSP70: role of toll-like receptor (TLR) 2 and TLR4. J Biol Chem 277: 15028–15034 - PubMed
-
- Brenner DA, O'Hara M, Angel P, Chojkier M, Karin M (1989) Prolonged activation of jun and collagenase genes by tumour necrosis factor-alpha. Nature 337: 661–663 - PubMed
-
- Bruhns P, Samuelsson A, Pollard JW, Ravetch JV (2003) Colony-stimulating factor-1-dependent macrophages are responsible for IVIG protection in antibody-induced autoimmune disease. Immunity 18: 573–581 - PubMed
-
- Calfon M, Zeng H, Urano F, Till JH, Hubbard SR, Harding HP, Clark SG, Ron D (2002) IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA. Nature 415: 92–96 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- DK083050/DK/NIDDK NIH HHS/United States
- P01 HL057346/HL/NHLBI NIH HHS/United States
- R01 AI079056/AI/NIAID NIH HHS/United States
- DP2 DK083050/DK/NIDDK NIH HHS/United States
- P30 ES006694/ES/NIEHS NIH HHS/United States
- R37 DK042394/DK/NIDDK NIH HHS/United States
- R01 DK090313/DK/NIDDK NIH HHS/United States
- R21 ES017829/ES/NIEHS NIH HHS/United States
- R01 CA154377/CA/NCI NIH HHS/United States
- R01 DK042394/DK/NIDDK NIH HHS/United States
- R56 AI079056/AI/NIAID NIH HHS/United States
- CA154377/CA/NCI NIH HHS/United States
- HL057346/HL/NHLBI NIH HHS/United States
- DK042394/DK/NIDDK NIH HHS/United States
- ES017829/ES/NIEHS NIH HHS/United States
- AI079056/AI/NIAID NIH HHS/United States
- DK090313/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
