

CAF-1 promotes Notch signaling through epigenetic control of target gene expression during Drosophila development
- PMID: 23942516
- PMCID: PMC3742146
- DOI: 10.1242/dev.094599
CAF-1 promotes Notch signaling through epigenetic control of target gene expression during Drosophila development
Abstract
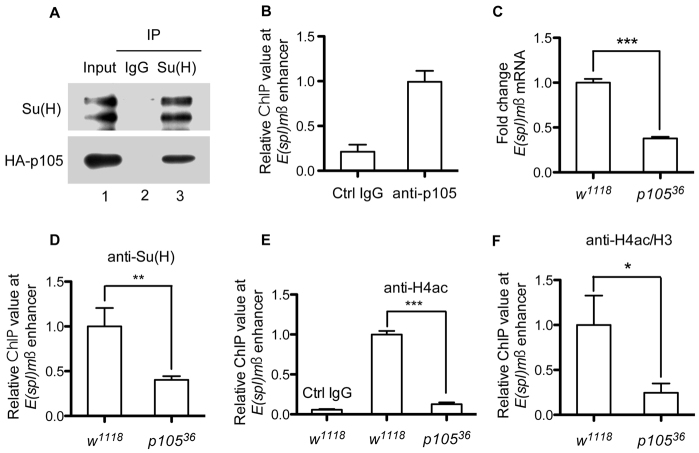
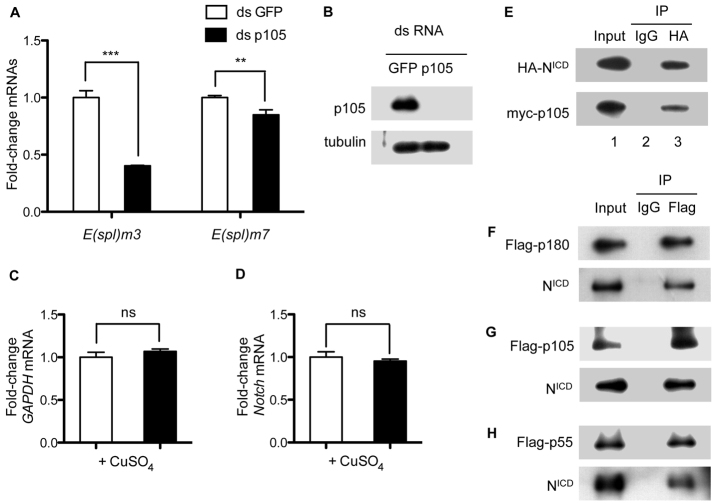
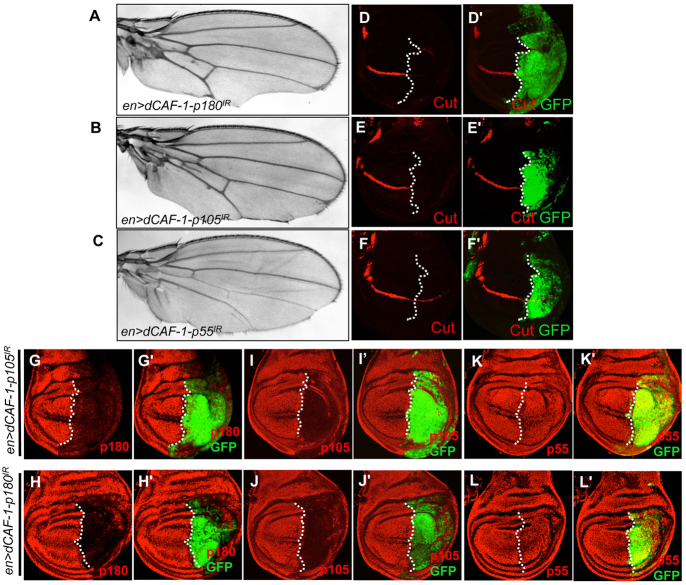
The histone chaperone CAF-1 is known for its role in DNA replication-coupled histone deposition. However, loss of function causes lethality only in higher multicellular organisms such as mice and flies, but not in unicellular organisms such as yeasts, suggesting that CAF-1 has other important functions than histone deposition during animal development. Emerging evidence indicates that CAF-1 also has a role in higher order chromatin organization and heterochromatin-mediated gene expression; it remains unclear whether CAF-1 has a role in specific signaling cascades to promote gene expression during development. Here, we report that knockdown of one of the subunits of Drosophila CAF-1, dCAF-1-p105 (Caf1-105), results in phenotypes that resemble those of, and are augmented synergistically by, mutations of Notch positive regulatory pathway components. Depletion of dCAF-1-p105 leads to abrogation of cut expression and to downregulation of other Notch target genes in wing imaginal discs. dCAF-1-p105 is associated with Suppressor of Hairless [Su(H)] and regulates its binding to the enhancer region of E(spl)mβ. The association of dCAF-1-p105 with Su(H) on chromatin establishes an active local chromatin status for transcription by maintaining a high level of histone H4 acetylation. In response to induced Notch activation, dCAF-1 associates with the Notch intracellular domain to activate the expression of Notch target genes in cultured S2 cells, manifesting the role of dCAF-1 in Notch signaling. Together, our results reveal a novel epigenetic function of dCAF-1 in promoting Notch pathway activity that regulates normal Drosophila development.
Keywords: CAF-1; Drosophila; Epigenetic regulation; H4 acetylation; Notch.
Figures
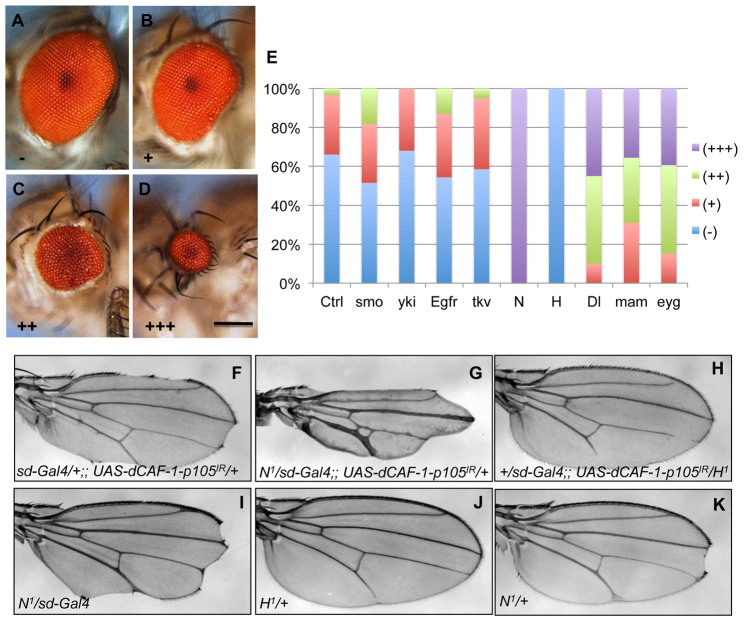
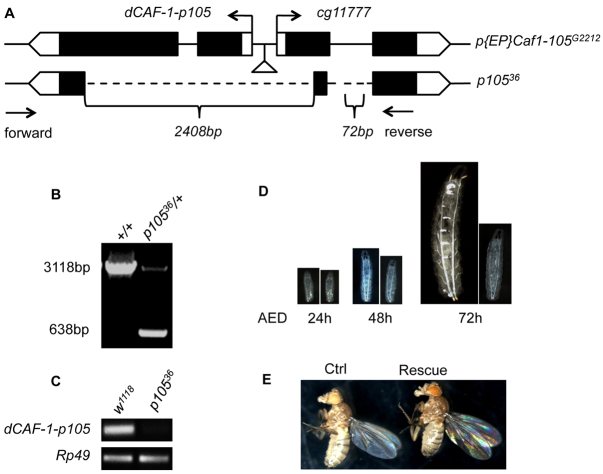
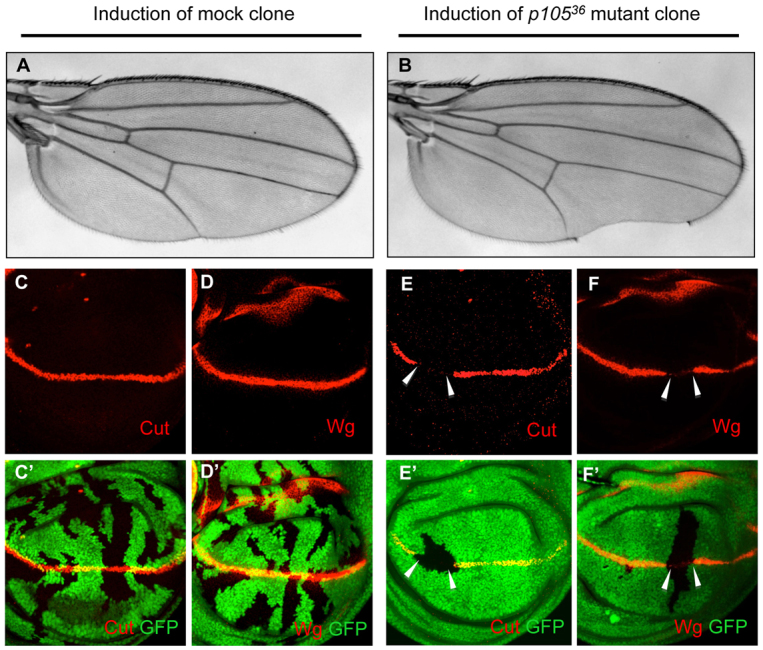
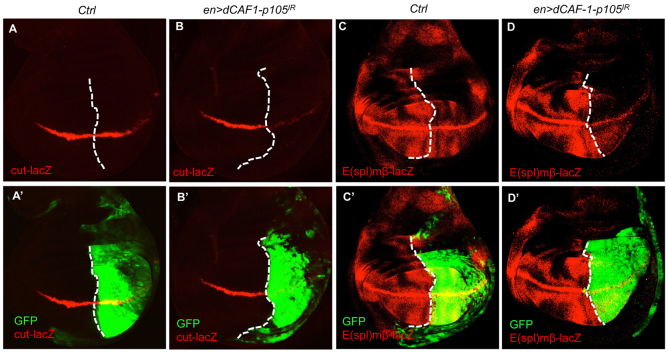
References
-
- Artavanis-Tsakonas S., Muskavitch M. A. (2010). Notch: the past, the present, and the future. Curr. Top. Dev. Biol. 92, 1–29 - PubMed
-
- Autran D., Baroux C., Raissig M. T., Lenormand T., Wittig M., Grob S., Steimer A., Barann M., Klostermeier U. C., Leblanc O., et al. (2011). Maternal epigenetic pathways control parental contributions to Arabidopsis early embryogenesis. Cell 145, 707–719 - PubMed
-
- Bray S. J. (2006). Notch signalling: a simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 7, 678–689 - PubMed
-
- Bray S., Musisi H., Bienz M. (2005). Bre1 is required for Notch signaling and histone modification. Dev. Cell 8, 279–286 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
