

Identification of biochemically distinct properties of the small ubiquitin-related modifier (SUMO) conjugation pathway in Plasmodium falciparum
- PMID: 23943616
- PMCID: PMC3784690
- DOI: 10.1074/jbc.M113.498410
Identification of biochemically distinct properties of the small ubiquitin-related modifier (SUMO) conjugation pathway in Plasmodium falciparum
Abstract
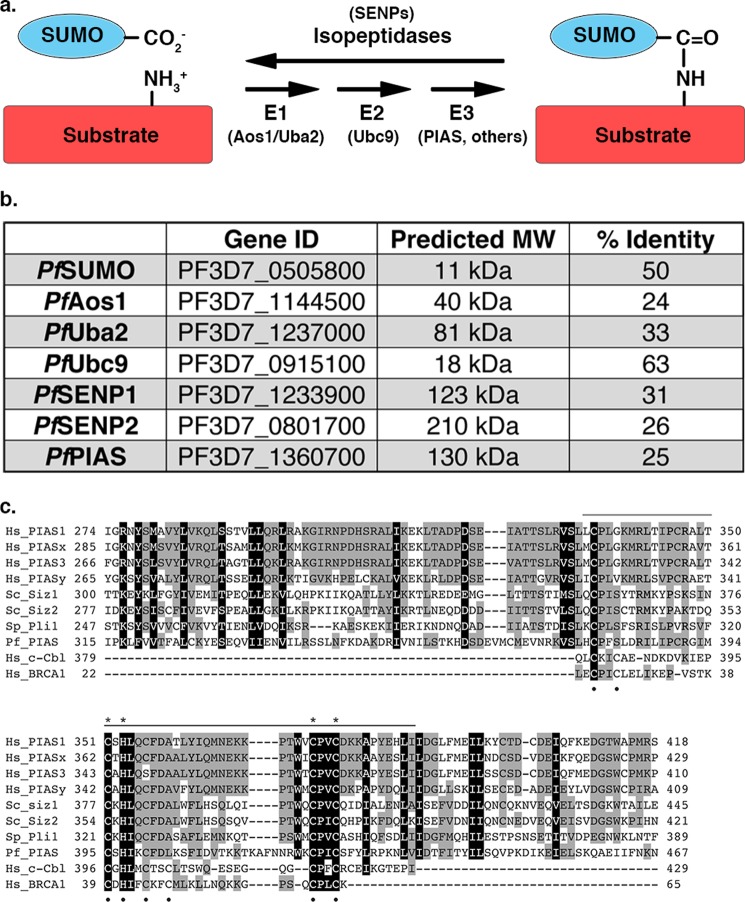
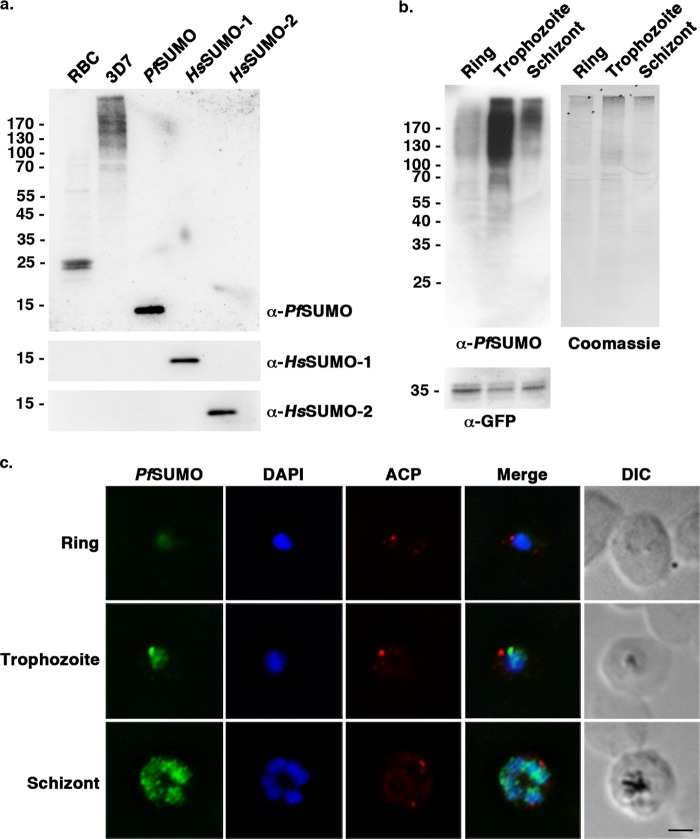
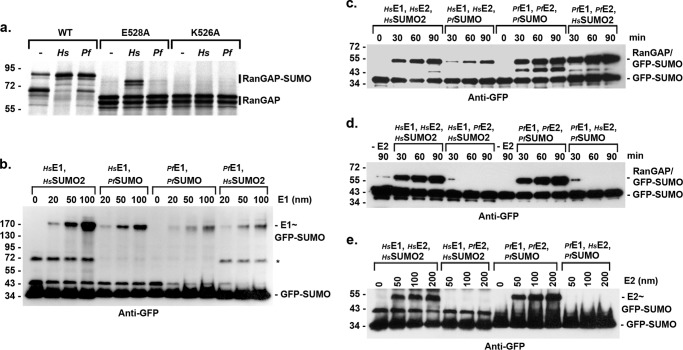
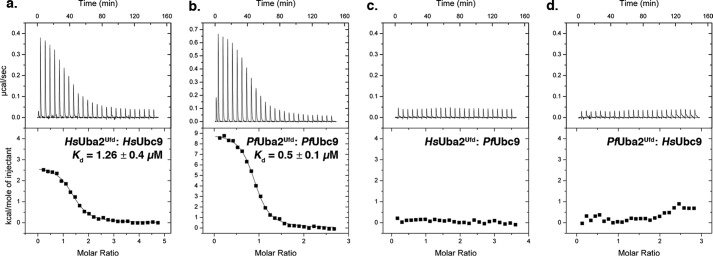
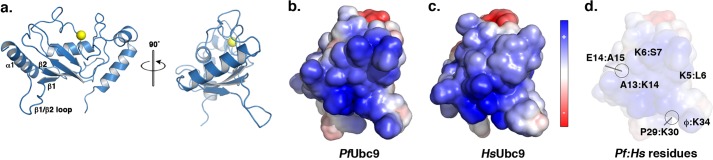
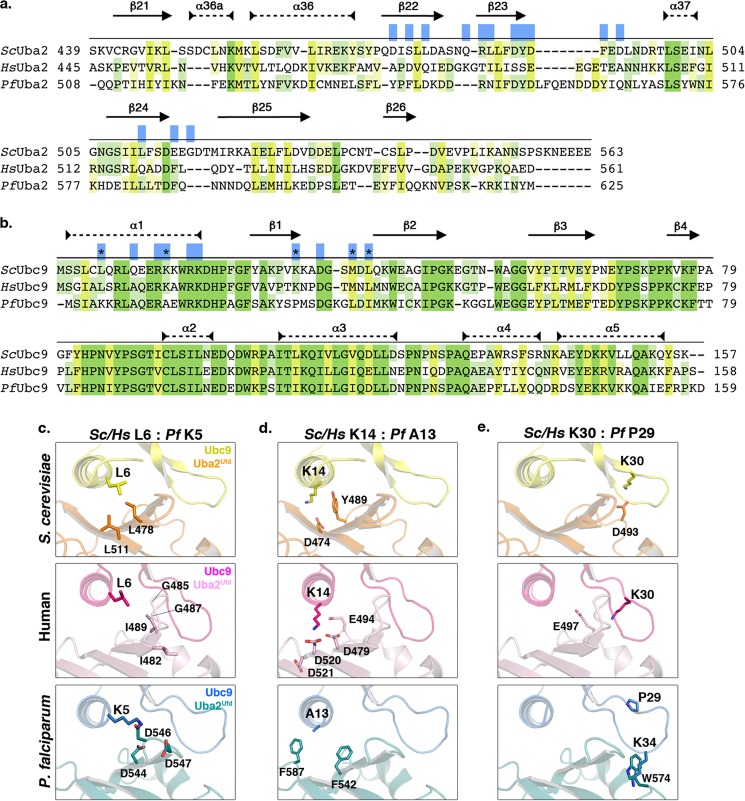
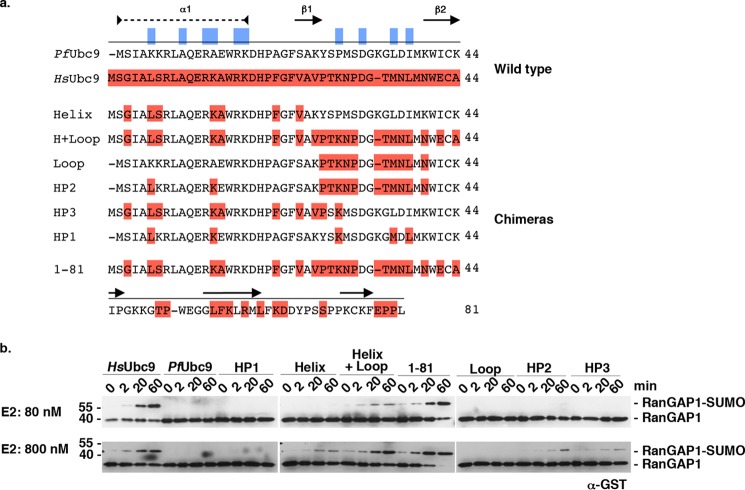
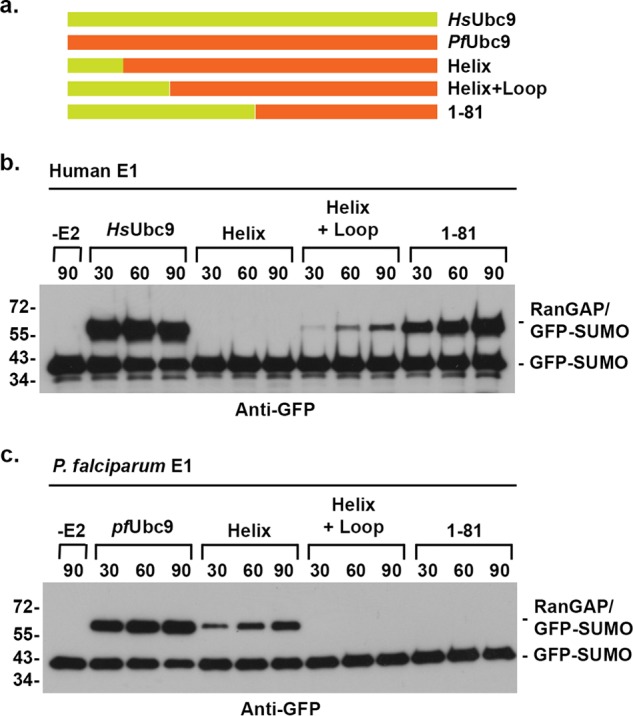
Small ubiquitin-related modifiers (SUMOs) are post-translationally conjugated to other proteins and are thereby essential regulators of a wide range of cellular processes. Sumoylation, and enzymes of the sumoylation pathway, are conserved in the malaria causing parasite, Plasmodium falciparum. However, the specific functions of sumoylation in P. falciparum, and the degree of functional conservation between enzymes of the human and P. falciparum sumoylation pathways, have not been characterized. Here, we demonstrate that sumoylation levels peak during midstages of the intra-erythrocyte developmental cycle, concomitant with hemoglobin consumption and elevated oxidative stress. In vitro studies revealed that P. falciparum E1- and E2-conjugating enzymes interact effectively to recognize and modify RanGAP1, a model mammalian SUMO substrate. However, in heterologous reactions, P. falciparum E1 and E2 enzymes failed to interact with cognate human E2 and E1 partners, respectively, to modify RanGAP1. Structural analysis, binding studies, and functional assays revealed divergent amino acid residues within the E1-E2 binding interface that define organism-specific enzyme interactions. Our studies identify sumoylation as a potentially important regulator of oxidative stress response during the P. falciparum intra-erythrocyte developmental cycle, and define E1 and E2 interactions as a promising target for development of parasite-specific inhibitors of sumoylation and parasite replication.
Keywords: Crystal Structure; Malaria; Plasmodium; Sumo; Sumoylation; Ubiquitin-conjugating Enzyme (Ubc).
Figures
Similar articles
-
Characterization and Structural Insights into Selective E1-E2 Interactions in the Human and Plasmodium falciparum SUMO Conjugation Systems.J Biol Chem. 2016 Feb 19;291(8):3860-70. doi: 10.1074/jbc.M115.680801. Epub 2015 Dec 23. J Biol Chem. 2016. PMID: 26697886 Free PMC article.
-
Biochemical characterization of SUMO-conjugating enzymes by in vitro sumoylation assays.Methods Enzymol. 2019;618:167-185. doi: 10.1016/bs.mie.2018.12.025. Epub 2019 Feb 11. Methods Enzymol. 2019. PMID: 30850051
-
Detection of SUMOylation in Plasmodium falciparum.Methods Mol Biol. 2016;1475:283-90. doi: 10.1007/978-1-4939-6358-4_19. Methods Mol Biol. 2016. PMID: 27631812
-
Protein interactions in the sumoylation cascade: lessons from X-ray structures.FEBS J. 2008 Jun;275(12):3003-15. doi: 10.1111/j.1742-4658.2008.06459.x. Epub 2008 May 17. FEBS J. 2008. PMID: 18492068 Review.
-
Molecular mechanisms in SUMO conjugation.Biochem Soc Trans. 2020 Feb 28;48(1):123-135. doi: 10.1042/BST20190357. Biochem Soc Trans. 2020. PMID: 31872228 Review.
Cited by
-
Implications of critical node-dependent unidirectional cross-talk of Plasmodium SUMO pathway proteins.Biophys J. 2022 Apr 19;121(8):1367-1380. doi: 10.1016/j.bpj.2022.03.022. Epub 2022 Mar 21. Biophys J. 2022. PMID: 35331687 Free PMC article.
-
Targeting SUMOylation in Plasmodium as a Potential Target for Malaria Therapy.Front Cell Infect Microbiol. 2021 Jun 10;11:685866. doi: 10.3389/fcimb.2021.685866. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34178724 Free PMC article. Review.
-
The ubiquitin system: an essential component to unlocking the secrets of malaria parasite biology.Mol Biosyst. 2014 Apr;10(4):715-23. doi: 10.1039/c3mb70506d. Epub 2014 Jan 30. Mol Biosyst. 2014. PMID: 24481176 Free PMC article. Review.
-
Ubiquitin-Like Modifiers: Emerging Regulators of Protozoan Parasites.Biomolecules. 2020 Oct 3;10(10):1403. doi: 10.3390/biom10101403. Biomolecules. 2020. PMID: 33022940 Free PMC article. Review.
-
Proteomic analysis of Plasmodium falciparum response to isocryptolepine derivative.PLoS One. 2019 Aug 8;14(8):e0220871. doi: 10.1371/journal.pone.0220871. eCollection 2019. PLoS One. 2019. PMID: 31393938 Free PMC article.
References
-
- Murray C. J., Rosenfeld L. C., Lim S. S., Andrews K. G., Foreman K. J., Haring D., Fullman N., Naghavi M., Lozano R., Lopez A. D. (2012) Global malaria mortality between 1980 and 2010. A systematic analysis. Lancet 379, 413–431 - PubMed
-
- Tilley L., Dixon M. W., Kirk K. (2011) The Plasmodium falciparum-infected red blood cell. Int. J. Biochem. Cell Biol. 43, 839–842 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
